
「節足動物」の版間の差分
| (3人の利用者による、間の6版が非表示) | |||
| 13行目: | 13行目: | ||
|上門階級なし = [[汎節足動物]] {{sname||Panarthropoda}} |
|上門階級なし = [[汎節足動物]] {{sname||Panarthropoda}} |
||
|門 = '''節足動物門''' {{sname||Arthropoda}} |
|門 = '''節足動物門''' {{sname||Arthropoda}} |
||
|学名='''{{sname|Arthropoda}}'''<br />{{AUY|[[カール・テオドール・フォン・シーボルト|von Siebold]]|1848}}<ref name="zhang2011">Zhi-Qiang Zhang (2011), |
|学名='''{{sname|Arthropoda}}'''<br />{{AUY|[[カール・テオドール・フォン・シーボルト|von Siebold]]|1848}}<ref name=":35" /><ref name="zhang2011">Zhi-Qiang Zhang (2011), “[https://doi.org/10.11646/zootaxa.3148.1.14 Phylum Arthropoda von Siebold, 1848]”. ''In'': Zhang Z.-Q. (Ed.), [http://www.mapress.com/zootaxa/list/2011/3148.html Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness]. ''Zootaxa'', Volume 3148, Magnolia Press, Pages 99-103.</ref> |
||
|和名 = 節足動物 |
|和名 = 節足動物 |
||
|英名 = Arthropod |
|英名 = Arthropod |
||
| 25行目: | 25行目: | ||
}} |
}} |
||
'''節足動物'''(せっそくどうぶつ、'''{{lang-en-short|Arthropod}}'''、[[学名]]:'''{{sname||Arthropoda}}''')とは、[[昆虫類]]・[[甲殻類]]・[[クモ]]類・[[ムカデ]]類など、[[外骨格]]と[[関節]]を持つ[[動物]]を含んだ[[タクソン|分類群]] |
'''節足動物'''(せっそくどうぶつ、'''{{lang-en-short|Arthropod}}'''、[[学名]]:'''{{sname||Arthropoda}}'''<ref name=":35">{{Cite book|title=Lehrbuch der vergleichenden Anatomie der Wirbellosen Thiere. Erster Theil. In Lehrbuch der vergleichenden Anatomie (eds C. T. von Siebold and H. Stannius)|url=https://www.biodiversitylibrary.org/bibliography/10707|publisher=Verlag von Veit & Comp.|date=1848|location=Berlin|first=C. Th. E. von|last=Siebold|first2=C. Th E. von|last2=Siebold|first3=Hermann|last3=Stannius|pages=679}}</ref>)とは、[[昆虫類]]・[[甲殻類]]・[[クモ]]類・[[ムカデ]]類など、[[外骨格]]と[[関節]]を持つ[[動物]]を含んだ[[タクソン|分類群]]。[[分類学]]上は'''節足動物門'''とされる。動物界最大かつ[[生物多様性|多様性]]の最も高い動物[[門 (分類学)|門]]であり<ref name=":4">{{Cite journal|last=Giribet|first=Gonzalo|last2=Edgecombe|first2=Gregory D.|date=2019-06-17|title=The Phylogeny and Evolutionary History of Arthropods|url=http://www.sciencedirect.com/science/article/pii/S0960982219304865|journal=Current Biology|volume=29|issue=12|pages=R592–R602|doi=10.1016/j.cub.2019.04.057|issn=0960-9822}}</ref><ref name="Daley2018" /><ref name=":1">{{Cite web|url=https://animaldiversity.org/accounts/Arthropoda/|title=Arthropoda (arthropods)|accessdate=2018-10-24|website=Animal Diversity Web|language=en}}</ref><ref name=":2">{{Cite book|title=The Arthropoda: A Phylogenetic Framework. In book: Arthropod Biology and Evolution – Molecules, Development, Morphology (pp.17-40)Chapter: The Arthropoda: a phylogenetic framework|last=Giribet|editor3-first=Giuseppe|editor2-last=Boxshall|editor2-first=Geoffrey|editor-last=Minelli|editor-first=Alessandro|last2=Edgecombe|first2=Gregory D.|first=Gonzalo|url=https://www.researchgate.net/publication/255686215|language=en|doi=10.1007/978-3-642-36160-9_2|pages=17–40|isbn=978-3-642-36159-3|location=Berlin, Heidelberg|date=2013|publisher=Springer Berlin Heidelberg|editor3-last=Fusco}}</ref>、現生[[種 (分類学)|種]]は全動物種の85%以上を占め、約110万種が記載される<ref>{{Cite book|和書 |title=節足動物の多様性と系統 |author=[[宮崎勝己]] |editor=[[石川良輔]]編、[[岩槻邦男]]・[[馬渡峻輔]]監修 |date=2008-04-11 |publisher=[[裳華房]] |chapter=節足動物における分類学の歴史 |pages=p.2 |series=バイオディバーシティ・シリーズ6 |id=ISBN 978-4-7853-5829-7}}</ref>。[[陸]]・[[海]]・[[空]]・[[土壌|土中]]・[[寄生]]などあらゆる場所に進出し、様々な[[生態系]]と深く関わっている。なお、いわゆる「[[虫]]」の範疇に入る動物は当動物門のものが多い<ref group="注釈">具体的には陸生を主体とする分類群([[多足類]]、[[六脚類]]、[[鋏角類]]の[[クモガタ類]]、[[甲殻類]]の[[等脚類]]など)が中心となり、また甲殻類の中で著しく特殊化した[[寄生]]性のものは虫と呼ばれるものが多い([[シタムシ]]、[[シダムシ]]、[[フクロムシ]]、[[ヒジキムシ]]など)。</ref>。 |
||
学名「{{sname|Arthropoda}}」は[[ギリシア語]]の「{{lang|grc|άρθρον}}」({{lang|grc-Latn|[[:wikt:arthron|arthron]]}}, 関節)と「{{lang|grc|πούς}}」({{lang|grc-Latn|[[:wikt:pous|pous]]}}, 脚)の合成語であり、本動物門の関節に分かれた[[付属肢]]([[関節肢]])に因んで名づけられた<ref name=":22">{{Cite book|title=A Dictionary of Entomology|url=https://books.google.com.tw/books?id=9IcmCeAjp6cC&pg=PA111&lpg=PA111&dq=Arthropoda+greek&source=bl&ots=Zp2OoiPS9p&sig=ACfU3U0SxrXnDKaFPzcM1qjiJ-FTvOn0PQ&hl=ja&sa=X&ved=2ahUKEwj9nN3pgY7kAhX_yYsBHf54DJ8Q6AEwEHoECAoQAQ#v=onepage&q=Arthropoda%20greek&f=false|publisher=CABI|date=2011|isbn=9781845935429|language=en}}</ref>。 |
学名「{{sname|Arthropoda}}」は[[ギリシア語]]の「{{lang|grc|άρθρον}}」({{lang|grc-Latn|[[:wikt:arthron|arthron]]}}, 関節)と「{{lang|grc|πούς}}」({{lang|grc-Latn|[[:wikt:pous|pous]]}}, 脚)の合成語であり、本動物門の関節に分かれた[[付属肢]]([[関節肢]])に因んで名づけられた<ref name=":22">{{Cite book|title=A Dictionary of Entomology|url=https://books.google.com.tw/books?id=9IcmCeAjp6cC&pg=PA111&lpg=PA111&dq=Arthropoda+greek&source=bl&ots=Zp2OoiPS9p&sig=ACfU3U0SxrXnDKaFPzcM1qjiJ-FTvOn0PQ&hl=ja&sa=X&ved=2ahUKEwj9nN3pgY7kAhX_yYsBHf54DJ8Q6AEwEHoECAoQAQ#v=onepage&q=Arthropoda%20greek&f=false|publisher=CABI|date=2011|isbn=9781845935429|language=en}}</ref>。 |
||
| 33行目: | 33行目: | ||
[[ファイル:Arthropod cuticle based upon Xvazquez edited to include english legend.PNG|160px|サムネイル|左|節足動物の[[クチクラ]]層の構造]] |
[[ファイル:Arthropod cuticle based upon Xvazquez edited to include english legend.PNG|160px|サムネイル|左|節足動物の[[クチクラ]]層の構造]] |
||
{{see also|関節肢|体節制}} |
{{see also|関節肢|体節制}} |
||
節足動物の形態は多様で、分類群によって様々な外見を持つ。体の表面は[[キチン質]]と[[タンパク質]]等からなる[[クチクラ]]([[:en:Arthropod cuticle|cuticle]])でできた[[外骨格]]([[:en:Exoskeleton|exoskeleton]])で覆われる。成長に伴い体のサイズが大きくなるときには、[[脱皮]]により古い外骨格は脱ぎ捨てられ、新しい外骨格が形成される。節足動物は'''[[体節制]]'''({{lang|en|[[:en:Segmentation (biology)|segmentation]]}})をも |
節足動物の形態は多様で、分類群によって様々な外見を持つ。体の表面は[[キチン質]]と[[タンパク質]]等からなる[[クチクラ]]([[:en:Arthropod cuticle|cuticle]])でできた[[外骨格]]([[:en:Exoskeleton|exoskeleton]])で覆われる。成長に伴い体のサイズが大きくなるときには、[[脱皮]]により古い外骨格は脱ぎ捨てられ、新しい外骨格が形成される。節足動物は'''[[体節制]]'''({{lang|en|[[:en:Segmentation (biology)|segmentation]]}})をもち、すなわち体は[[体節]](somite)という単位の繰り返し構造からなり、各体節は原則として1対の[[付属肢]]をもつ([[節足動物#付属肢|後述]])<ref name=":19">{{Cite book|title=Arthropod Biology and Evolution: Molecules, Development, Morphology|last=Fusco|editor3-first=Giuseppe|editor2-last=Boxshall|editor2-first=Geoffrey|editor-last=Minelli|editor-first=Alessandro|last2=Minelli|first2=Alessandro|first=Giuseppe|url=https://doi.org/10.1007/978-3-642-36160-9_9|language=en|doi=10.1007/978-3-642-36160-9_9|pages=197–221|isbn=978-3-642-36160-9|location=Berlin, Heidelberg|date=2013|publisher=Springer|editor3-last=Fusco}}</ref>。体の背腹は、それぞれ[[背板]](tergite, [[:en:Tergum|tergum]])と[[腹板]](sternite, [[:en:Sternum (arthropod anatomy)|sternum]])という外骨格に覆われており、体節の両側で更に側板([[:en:Pleuron (insect anatomy)|pleuron]])を持つ場合もある。表面を被うこれらの外骨格も体節単位になっており、体節の間は関節状に可動であることが多い。体の先頭の体節は[[先節]](ocular somite、古典的には[[口前葉]] acron)といい、節足動物の[[眼]]と[[口]]はここに由来する<ref name="head" />。一部の分類群は、体の末端に[[尾節]]({{lang|en|[[:en:telson|telson]]}})という[[尾]]のような構造もある<ref name=":19" />。 |
||
<gallery mode="packed" heights="200px"> |
<gallery mode="packed" heights="200px"> |
||
Tagma.png|[[昆虫]]の体制:頭部・胸部・腹部からなる |
Tagma.png|[[昆虫]]の体制:頭部・胸部・腹部からなる |
||
| 39行目: | 39行目: | ||
Archindae characters.jpg|[[クモ綱|クモガタ類]]の体制:前体([[頭胸部]])・後体(腹部)からなる |
Archindae characters.jpg|[[クモ綱|クモガタ類]]の体制:前体([[頭胸部]])・後体(腹部)からなる |
||
</gallery> |
</gallery> |
||
ただし、節足動物は'''異規体節制'''({{lang|en|Heteronomous metamerism}}<ref>{{Cite book|title=Biology of Arthropoda|url=https://books.google.com.tw/books?id=Hd4OEDo4gbwC&pg=PA317&lpg=PA317&dq=Heteronomous+metamerism&source=bl&ots=C-fPzbQ6yq&sig=ACfU3U0CEtIzRio5BqsshrDHKwht9xkVVA&hl=ja&sa=X&ved=2ahUKEwiL-9ryqI_kAhVpwosBHbWrBz8Q6AEwAnoECAYQAQ#v=onepage&q=Heteronomous%20metamerism&f=false|publisher=Discovery Publishing House|date=2004|isbn=9788171418978|language=en|first=D. R.|last=Khanna}}</ref>)がある程度以上発達し、複数の体節が組み合わされ、外観上あるいは機能上の単位を構成する({{lang|en|tagmosis}}、{{lang|en|tagmatization}})<ref name=":19" />例が多く見られる。例えば、体を「[[頭部]]・[[胸部]]・[[腹部]]」「頭部・胸部・尾部」「前体・中体・終体」などの3部、または「頭部・胴部」「[[頭胸部]]・腹部」「前体・後体」などの2部に分けて呼ぶ場合があり、これは節足動物の各分類群ごとの特徴として用いられる<ref name=":19" />。 |
ただし、節足動物は'''異規体節制'''({{lang|en|Heteronomous metamerism}}<ref>{{Cite book|title=Biology of Arthropoda|url=https://books.google.com.tw/books?id=Hd4OEDo4gbwC&pg=PA317&lpg=PA317&dq=Heteronomous+metamerism&source=bl&ots=C-fPzbQ6yq&sig=ACfU3U0CEtIzRio5BqsshrDHKwht9xkVVA&hl=ja&sa=X&ved=2ahUKEwiL-9ryqI_kAhVpwosBHbWrBz8Q6AEwAnoECAYQAQ#v=onepage&q=Heteronomous%20metamerism&f=false|publisher=Discovery Publishing House|date=2004|isbn=9788171418978|language=en|first=D. R.|last=Khanna}}</ref>)がある程度以上発達し、複数の体節が組み合わされ、外観上あるいは機能上の単位を構成する({{lang|en|tagmosis}}、{{lang|en|tagmatization}})<ref name=":19" />例が多く見られる。例えば、体を「[[頭部]]・[[胸部]]・[[腹部]]」「頭部・胸部・尾部」「前体・中体・終体」などの3部、または「頭部・胴部」「[[頭胸部]]・腹部」「前体・後体」などの2部に分けて呼ぶ場合があり、これは節足動物の各分類群ごとの特徴として用いられる<ref name=":19" />。特に前方の合体節(頭部・頭胸部・前体)は往々にして体節の癒合が進み、外見上では元の体節構造が見当たらず、すべて単一の外骨格に覆われている<ref name=":19" />。 |
||
なお、一部の寄生性の分類群は |
なお、一部の寄生性の分類群には極端に特化し、外見上の体節構造が全く見当たらない例もある<ref name=":19" />。 |
||
=== 付属肢 === |
=== 付属肢 === |
||
| 47行目: | 47行目: | ||
[[ファイル:20200213 arthropod uniramus arthropodized appendage morphology ja.png|350px|サムネイル|[[関節肢]]の基本外部形態]] |
[[ファイル:20200213 arthropod uniramus arthropodized appendage morphology ja.png|350px|サムネイル|[[関節肢]]の基本外部形態]] |
||
[[ファイル:Evolution insect mouthparts coloured.png|220px|サムネイル|右|様々な[[昆虫]]の口器( a:[[触角]]、c:[[複眼]]、lb:[[上唇 (節足動物)|上唇]]、lr:[[下唇#六脚類|下唇]]、md:[[大顎]]、mx:[[小顎]])]] |
[[ファイル:Evolution insect mouthparts coloured.png|220px|サムネイル|右|様々な[[昆虫]]の口器( a:[[触角]]、c:[[複眼]]、lb:[[上唇 (節足動物)|上唇]]、lr:[[下唇#六脚類|下唇]]、md:[[大顎]]、mx:[[小顎]])]] |
||
各体節からは、原則として1対の'''[[関節肢]]'''({{lang|en|[[:en:arthropod leg|arthropodized appendage]]}})という[[付属肢]]が出ている。これが「節足動物」という名前およびその学名の由来となっている<ref name=":22" />。関節肢も体と同様に外骨格で覆われ、[[関節]]によって分かれた肢節(podomere)からなる。関節肢は分類群や |
各体節からは、原則として1対の'''[[関節肢]]'''({{lang|en|[[:en:arthropod leg|arthropodized appendage]]}})という本群に特有の[[付属肢]]が出ている。これが「節足動物」という名前およびその学名の由来となっている<ref name=":22" />。関節肢も体と同様に外骨格で覆われ、[[関節]]によって分かれた肢節(podomere)からなる。関節肢は所属する分類群や位置によって歩脚・[[遊泳]]脚・[[はさみ (動物)|鋏]]・[[はさみ (動物)#亜鋏状の構造|鎌]]・[[顎]]・[[触角]]・[[鰓]]・[[生殖肢]]など様々な機能に応じて色んな形に特化している<ref name=":19" />。例えば頭部には感覚用の触角と摂食用の顎、胴部には移動用の歩脚を持つなど、節足動物は、往々にして異なる機能を担った様々な関節肢を掛け合わせて、「[[アーミーナイフ]]のように、独自の機能を有する複数の道具が同時にセットされる」とも比喩される<ref>Ruppert, E. E.; R. S. Fox; R. D. Barnes (2004), Invertebrate Zoology (7th ed.), Brooks/Cole, ISBN 0-03-025982-7</ref>。また、節足動物は多くが口の直前に[[上唇 (節足動物)|上唇]](labrum)もしくは[[ハイポストーマ]](hypostome)と呼ばれる1枚の蓋状の構造体があり、これも付属肢由来の部分ではないかと思われる<ref name=":28" /><ref name=":29" /><ref name="stem" /><ref name="head" />。なお、前述の体節のように、寄生性[[甲殻類]]や[[昆虫]]の[[幼虫]]の中には、関節肢が不明瞭もしくは完全に退化消失した例もある<ref name=":19" />。 |
||
<gallery mode="packed" heights="150px"> |
<gallery mode="packed" heights="150px"> |
||
General malacostracan en.svg|[[軟甲類]]の体制模式図:頭部には2対の触角と3対の顎・胸部には8対の胸肢・腹部には5対の腹肢と1対の尾肢をもつ |
General malacostracan en.svg|[[軟甲類]]の体制模式図:頭部には2対の触角と3対の顎・胸部には8対の胸肢・腹部には5対の腹肢と1対の尾肢をもつ |
||
FMIB 46766 Lupee pelagienne.jpeg|鋏型の第1脚、歩脚型の第2-4脚とヘラ状の第5脚(遊泳脚)をもつ[[タイワンガザミ]](ワタリガニ) |
FMIB 46766 Lupee pelagienne.jpeg|鋏型の第1脚、歩脚型の第2-4脚とヘラ状の第5脚(遊泳脚)をもつ[[タイワンガザミ]](ワタリガニ) |
||
</gallery> |
</gallery> |
||
| ⚫ | |||
[[ファイル:20211214 arthropod biramous arthropodized appendage morphology ja.png|サムネイル|二叉型付属肢の基本外部形態]] |
|||
関節肢は、内肢({{lang|en|endopod}})と外肢({{lang|en|exopod}})もしくは外葉({{lang|en|exite}})をもつ'''二叉型付属肢'''(二肢型付属肢、{{lang|en|biramous appendage}})が[[共有原始形質|祖先形質]]であると思われる。しかし、現生節足動物の中にこの性質を明瞭に受け継いだのは多くが[[甲殻類]]で、他の分類群においては内肢しか見られない'''単枝型付属肢'''(単肢型付属肢、{{lang|en|uniramous appendage}})がほとんどである。また、[[三葉虫]]を含んだ[[:en:Artiopoda|Artiopoda]]類など、多くの化石節足動物の分類群は胴部の付属肢がヒレ状の外肢/外葉と歩脚状の内肢からなる二叉型である。 |
|||
関節肢は、内肢({{lang|en|endopod}})と外肢({{lang|en|exopod}})もしくは外葉({{lang|en|exite}}、副肢 epipod)をもつ'''二叉型付属肢'''(二肢型付属肢、{{lang|en|biramous appendage}})が基本だと思われる<ref name="Alacaris" /><ref name=":36">{{Cite journal|last=Liu|first=Yu|last2=Edgecombe|first2=Gregory D.|last3=Schmidt|first3=Michel|last4=Bond|first4=Andrew D.|last5=Melzer|first5=Roland R.|last6=Zhai|first6=Dayou|last7=Mai|first7=Huijuan|last8=Zhang|first8=Maoyin|last9=Hou|first9=Xianguang|date=2021-07-30|title=Exites in Cambrian arthropods and homology of arthropod limb branches|url=https://www.nature.com/articles/s41467-021-24918-8|journal=Nature Communications|volume=12|issue=1|pages=4619|language=en|doi=10.1038/s41467-021-24918-8|issn=2041-1723}}</ref>。しかし、現生節足動物の中でこの性質を明瞭に受け継いだのは多くが[[甲殻類]]で<ref name=":37">{{Cite web|title=Exopodites , Epipodites and Gills in Crustaceans|url=https://www.semanticscholar.org/paper/Exopodites-%2C-Epipodites-and-Gills-in-Crustaceans-Boxshall-Jaume/bca391fcdd95b5c4d403bca43925b2fe28b65331|website=www.semanticscholar.org|date=2009|accessdate=2021-12-23|language=en|first=G.|last=Boxshall}}</ref>、他の分類群においては内肢しか見られない'''単枝型付属肢'''(単肢型付属肢、{{lang|en|uniramous appendage}})がほとんどである。また、[[三葉虫]]と[[メガケイラ類]]をはじめとする多くの[[古生代]]の絶滅分類群は、胴部の付属肢がヒレ状の外肢と歩脚状の内肢でできた二叉型である<ref name="Alacaris" /><ref name=":36" />。 |
|||
<gallery> |
<gallery> |
||
| ⚫ | |||
CrustaceaAppendages.png|様々な[[甲殻類]]の二叉型附属肢(en:内肢、ex:外肢、ep:外葉) |
CrustaceaAppendages.png|様々な[[甲殻類]]の二叉型附属肢(en:内肢、ex:外肢、ep:外葉) |
||
Gnathopods nl.png|鋏状(右)と亜鋏状(左)の[[関節肢]] |
Gnathopods nl.png|鋏状(右)と亜鋏状(左)の[[関節肢]] |
||
| 64行目: | 66行目: | ||
=== 運動 === |
=== 運動 === |
||
{{see also|関節肢}} |
{{see also|関節肢}} |
||
関節の隙間は |
関節の隙間は柔らかい'''節間膜'''({{lang|en|arthrodial membrane}})に覆われており、2つの関節の外骨格内側に付着する[[筋肉]]を介して運動をする。胴部の関節は様々な方向へ湾曲できることが多いが、[[関節肢]]の関節は往々にして1対の蝶番のような構造、いわゆる'''関節丘'''(ピボット、{{lang|en|pivot}})に固定されて一つの平面上でしか動かない。そのため節足動物の関節肢、特に基部は往々にして複数の節に分かれて、様々な動きに対応できるようになる<ref>{{Cite journal|last=Turetzek|first=Natascha|last2=Pechmann|first2=Matthias|last3=Schomburg|first3=Christoph|last4=Schneider|first4=Julia|last5=Prpic|first5=Nikola-Michael|date=2016-01-01|title=Neofunctionalization of a Duplicate dachshund Gene Underlies the Evolution of a Novel Leg Segment in Arachnids|url=https://doi.org/10.1093/molbev/msv200|journal=Molecular Biology and Evolution|volume=33|issue=1|pages=109–121|doi=10.1093/molbev/msv200|issn=0737-4038}}</ref>。また、外骨格は体の内側へ延長し、いわゆる'''[[腱]]'''(内突起、{{Lang|en|internal tendon}}, {{lang|en|apodeme}})となり、筋肉に付着面を提供することが普通である<ref>{{Cite web|title=Appendage - an overview {{!}} ScienceDirect Topics|url=https://www.sciencedirect.com/topics/agricultural-and-biological-sciences/appendage|website=www.sciencedirect.com|accessdate=2021-05-24}}</ref>。例えば[[カニ]]の[[はさみ (動物)|鋏]]の中では、可動指の関節に繋がった、大量の筋肉が付着する板状の腱が見られる。また、筋肉の代わりに体中の血リンパ(後述)の[[水圧]]を調整することで関節肢を動かす例もある<ref name=":33">Rainer F. Foelix (2011). [https://www.academia.edu/13113405/Biology_of_Spiders ''Biology of spiders'' (3rd ed.)]. New York: Oxford University Press. [[ISBN]] [[特別:文献資料/978-0-19-981324-7|978-0-19-981324-7]]. [[Online Computer Library Center|OCLC]] 693776865</ref>。 |
||
=== 神経系と循環系 === |
=== 神経系と循環系 === |
||
| 70行目: | 72行目: | ||
[[File:Insects nervous system ja.png|サムネイル|238px|[[昆虫]]の[[神経系]]模式図<hr />P: 前大脳、D: 中大脳、T: 後大脳、CV: 腹髄神経索]] |
[[File:Insects nervous system ja.png|サムネイル|238px|[[昆虫]]の[[神経系]]模式図<hr />P: 前大脳、D: 中大脳、T: 後大脳、CV: 腹髄神経索]] |
||
{{see also|はしご形神経系}} |
{{see also|はしご形神経系}} |
||
多くの[[左右相称動物]]と同様、節足動物は[[体腔]]を持ち、[[消化管]]は体の前後に貫通し、いわゆる[[口]]と[[肛門]]という2つ開 |
多くの[[左右相称動物]]と同様、節足動物は[[体腔]]を持ち、[[消化管]]は体の前後に貫通し、いわゆる[[口]]と[[肛門]]という2つ開口を持つ。体節に貫通する紐状の[[心臓]]と[[中枢神経]]はそれぞれ体の背面と腹面に付く。 |
||
| ⚫ | |||
[[体節制]]をもつ他の[[前口動物]]に似て、節足動物の[[神経系]]の様式は'''[[はしご形神経系]]'''({{lang|en|ladder-like nervous system}})である。腹側(ただし脳は背側<ref name="Daley2018" />)の1対の神経索が体の前後を走り、各[[体節]]に由来する[[神経節]]({{lang|en|ganglion}})は、左右の連絡(横連合、{{lang|en|commissure}})で繋がっている。 |
|||
| ⚫ | |||
体の前端部には[[脳]]があり、[[食道]]の前上方にあることから[[食道上神経節]](supraoesophageal ganglion、大脳神経節<ref name="murakami">{{Cite book|和書|author=村上安則|editor=公益社団法人 日本動物学会|chapter=神経系の発生 {{small|―ひとりでに出来上がるコンピュータ}}|title=動物学の百科事典|date=2018-09-28|publisher=丸善出版|pages=310-311|isbn=978-4621303092}}</ref>)とも呼ぶ。この脳は先頭3つの体節([[先節]]・第1体節・第2体節)と共に3対の[[神経節]]が融合した結果であり、前大脳({{lang|en|protocerebrum}})・中大脳({{lang|en|deutocerebrum}})・後大脳({{lang|en|tritocerebrum}})という3つの脳神経節から構成される<ref name="mizunami" /><ref name="stem" /><ref name="head" />。前大脳には複眼からの[[視覚]]情報を処理する[[視葉]](optic lobe)、[[嗅覚]]の識別や記憶および感覚神経の統御を司る[[キノコ体]]({{lang|en|[[:en:mushroom body|mushroom body]]}})、視覚行動の統御を行う[[中心複合体]](central body)を持つ<ref name="mizunami" />。脳は前大脳をはじめとして背側にあるため、中央もしくは直後から食道を囲み、[[食道神経環]]([[:en:Circumesophageal nerve ring|circumesophageal nerve ring]])を介して腹側の腹神経索([[:en:Ventral nerve cord|ventral nerve cord]])に連結する。[[昆虫]]・[[甲殻類]]などの[[大顎類]]の場合、食道神経環の直後は[[顎]]([[大顎]]・[[小顎]]/[[下唇 (節足動物)|下唇]])に対応する神経節で、まとめて[[食道下神経節]](suboesophageal ganglion、顎神経節<ref name="murakami" />)といい<ref name="mizunami">{{Cite book|和書|author=水波誠|editor=公益社団法人 日本動物学会|chapter=昆虫の微小脳 {{small|―小さな脳のすごい働き}}|title=動物学の百科事典|date=2018-09-28|publisher=丸善出版|pages=362-363|isbn=978-4621303092}}</ref>、[[ハエ]]や[[ハチ]]、[[チョウ]]などにおいては脳と融合し[[頭部神経節]]を構成する(この場合は食道上神経節のみならず、食道下神経節も脳の一部と扱う)<ref name="mizunami" />。一部の体節の融合が進み、神経が集中してはしご形が不明瞭な場合もあり、例えば[[カブトガニ類|カブトガニ]]や[[クモガタ類]]の前体において脳と腹神経索を集約させた synganglion、および[[カニ]]や派生的な[[昆虫]]において著しく集約した胸部と腹部の神経節がその例である<ref name=":20">{{Cite journal|last=Smarandache-Wellmann|first=Carmen Ramona|date=10 24, 2016|title=Arthropod neurons and nervous system|url=https://www.ncbi.nlm.nih.gov/pubmed/27780069|journal=Current biology: CB|volume=26|issue=20|pages=R960–R965|doi=10.1016/j.cub.2016.07.063|issn=1879-0445|pmid=27780069}}</ref>。 |
|||
{{-}} |
{{-}} |
||
| 80行目: | 84行目: | ||
[[ファイル:Insect antennae.svg|330px|サムネイル|右|様々な[[昆虫]]の[[触角]]]] |
[[ファイル:Insect antennae.svg|330px|サムネイル|右|様々な[[昆虫]]の[[触角]]]] |
||
{{see also|触角}} |
{{see also|触角}} |
||
節足動物は様々な[[感覚器]]を通じて周りの環境を感知する。体表は常に剛毛(感覚毛、[[:en:Setae|setae]])をもち、種によっては[[触覚]]・[[振動]]・[[音]]・[[水流]]や[[気流]]・[[温度]]・[[匂い]]や[[味]]・[[化学物質]]など視力以外の感覚を持つ。[[鋏角類]]以外の節足動物の頭部は、往々にして'''[[触角]]'''({{lang|en|[[:w:Antenna|Antenna]]}})という[[関節肢]]をもち、ほとんどの場合は重要な感覚器官である。なお、触角のない[[鋏角類]]の中でも、[[ウデムシ]]や[[サソリモドキ]]の様に一部の歩脚が感覚器官に特化した例がある<ref name=":5">{{Cite journal|last=Lamsdell|first=James C.|last2=Dunlop|first2=Jason A.|title=Segmentation and tagmosis in Chelicerata|url=https://www.academia.edu/28212892/Segmentation_and_tagmosis_in_Chelicerata|journal=Arthropod Structure & Development|volume=46|issue=3|pages=395–418|language=en|issn=1467-8039}}</ref>。 |
節足動物は様々な[[感覚器]]を通じて周りの環境を感知する。体表は常に剛毛(刺毛、感覚毛、[[:en:Setae|setae]])をもち、種によっては[[触覚]]・[[振動]]・[[音]]・[[水流]]や[[気流]]・[[温度]]・[[匂い]]や[[味]]・[[化学物質]]など視力以外の感覚を持つ。[[鋏角類]]以外の節足動物の頭部は、往々にして'''[[触角]]'''({{lang|en|[[:w:Antenna|Antenna]]}})という[[関節肢]]をもち、ほとんどの場合は重要な感覚器官である。なお、触角のない[[鋏角類]]の中でも、[[ウデムシ]]や[[サソリモドキ]]の様に一部の歩脚が感覚器官に特化した例がある<ref name=":5">{{Cite journal|last=Lamsdell|first=James C.|last2=Dunlop|first2=Jason A.|title=Segmentation and tagmosis in Chelicerata|url=https://www.academia.edu/28212892/Segmentation_and_tagmosis_in_Chelicerata|journal=Arthropod Structure & Development|volume=46|issue=3|pages=395–418|language=en|issn=1467-8039}}</ref>。 |
||
[[ファイル:Orthoptera mouthparts SEM.jpg|220px|サムネイル|右|[[バッタ]]の[[小顎]]髭(d)と[[小顎#六脚類|下唇]]鬚(e)]] |
[[ファイル:Orthoptera mouthparts SEM.jpg|220px|サムネイル|右|[[バッタ]]の[[小顎]]髭(d)と[[小顎#六脚類|下唇]]鬚(e)]] |
||
他にも[[昆虫]]の小顎と下唇にある顎鬚(palp)が[[嗅覚]]や[[味覚]]に関わり、[[サソリ]]の櫛状板・[[ヒヨケムシ]]のラケット器官・一部の[[昆虫]]と[[甲殻類]]の後端にある[[尾毛]]([[:en:cercus|cercus]]、[[:en:caudal ramus|caudal ramus]])が感覚器官として用いられるほか、[[コオロギ]]や[[キリギリス]]が前脚に<ref>{{Cite journal|date=1993-04-01|title=The tettigoniid (Orthoptera : Tettigoniidae) ear: Multiple functions and structural diversity|url=https://www.sciencedirect.com/science/article/pii/002073229390009P|journal=International Journal of Insect Morphology and Embryology|volume=22|issue=2-4|pages=185–205|language=en|doi=10.1016/0020-7322(93)90009-P|issn=0020-7322}}</ref>・[[バッタ]]が腹部に<ref>{{Cite web|url=https://www.archie-west.ac.uk/projects/systems-biology/acoustic-receivers-from-insect-ear-to-next-generation-sensor/|title=Acoustic Receivers: From Insect Ear to Next-Generation Sensor {{!}} ARCHIE-WeSt|accessdate=2018年12月20日|publisher=}}</ref>・[[カマキリ]]が後胸部腹面<ref>{{Cite journal|last=Yager|first=David|last2=Hoy|first2=Ron|date=1988-01-01|title=The midline metathoracic ear of the praying mantis, Mantis religiosa|url=https://www.researchgate.net/publication/19470569_The_midline_metathoracic_ear_of_the_praying_mantis_Mantis_religiosa|journal=Cell and tissue research|volume=250|pages=531-541|doi=10.1007/BF00218944}}</ref>に有する、特殊化した聴覚器官である[[鼓膜器官]]が知られる。 |
他にも[[昆虫]]の小顎と下唇にある顎鬚(palp)が[[嗅覚]]や[[味覚]]に関わり、[[サソリ]]の櫛状板・[[ヒヨケムシ]]のラケット器官・一部の[[昆虫]]と[[甲殻類]]の後端にある[[尾毛]]([[:en:cercus|cercus]]、[[:en:caudal ramus|caudal ramus]])が感覚器官として用いられるほか、[[コオロギ]]や[[キリギリス]]が前脚に<ref>{{Cite journal|date=1993-04-01|title=The tettigoniid (Orthoptera : Tettigoniidae) ear: Multiple functions and structural diversity|url=https://www.sciencedirect.com/science/article/pii/002073229390009P|journal=International Journal of Insect Morphology and Embryology|volume=22|issue=2-4|pages=185–205|language=en|doi=10.1016/0020-7322(93)90009-P|issn=0020-7322}}</ref>・[[バッタ]]が腹部に<ref>{{Cite web|url=https://www.archie-west.ac.uk/projects/systems-biology/acoustic-receivers-from-insect-ear-to-next-generation-sensor/|title=Acoustic Receivers: From Insect Ear to Next-Generation Sensor {{!}} ARCHIE-WeSt|accessdate=2018年12月20日|publisher=}}</ref>・[[カマキリ]]が後胸部腹面<ref>{{Cite journal|last=Yager|first=David|last2=Hoy|first2=Ron|date=1988-01-01|title=The midline metathoracic ear of the praying mantis, Mantis religiosa|url=https://www.researchgate.net/publication/19470569_The_midline_metathoracic_ear_of_the_praying_mantis_Mantis_religiosa|journal=Cell and tissue research|volume=250|pages=531-541|doi=10.1007/BF00218944}}</ref>に有する、特殊化した聴覚器官である[[鼓膜器官]]が知られる。 |
||
| 94行目: | 98行目: | ||
[[ファイル:FLY EYE.jpg|220px|サムネイル|左|[[ハエ]]の[[複眼]]のクローズアップ([[走査型電子顕微鏡]]写真)]] |
[[ファイル:FLY EYE.jpg|220px|サムネイル|左|[[ハエ]]の[[複眼]]のクローズアップ([[走査型電子顕微鏡]]写真)]] |
||
[[ファイル:Eyes and ocelli of an orchid bee (Apidae, Euglossa hansoni (Moure)) (36522833083).jpg|サムネイル|220px|右|[[ハチ]]の3つの単眼(中央)と2つの複眼(左右)]] |
[[ファイル:Eyes and ocelli of an orchid bee (Apidae, Euglossa hansoni (Moure)) (36522833083).jpg|サムネイル|220px|右|[[ハチ]]の3つの単眼(中央)と2つの複眼(左右)]] |
||
節足動物は、'''中眼'''({{lang|en|median eye}})と'''側眼'''({{lang|en|lateral eye}})という[[先節]]に由来する<ref name="head" />2種類の[[眼]]を持ち、その中で中眼は'''[[複眼と単眼|単眼]]'''({{lang|la|ocellus}}, {{lang|en|simple eye}})で、側眼は'''[[複眼]]'''(compound eye)であることが基本と思われる<ref name=":21" />。しかしその片方しか持たず、複眼が単眼(側単眼)と化したり、更に祖先形質として眼をもつ多くの動物と同様、眼が二次的に退化消失した節足動物もある。 |
節足動物は、'''中眼'''({{lang|en|median eye}})と'''側眼'''({{lang|en|lateral eye}})という[[先節]]に由来する<ref name="head" />2種類の[[眼]]を持ち、その中で中眼は'''[[複眼と単眼|単眼]]'''({{lang|la|ocellus}}, {{lang|en|simple eye}})で、側眼は'''[[複眼]]'''(compound eye)であることが基本と思われる<ref name=":21" /><ref name=":38" />。しかしその片方しか持たず、複眼が単眼(側単眼)と化したり、更に祖先形質として眼をもつ多くの動物の分類群と同様、眼が二次的に退化消失した節足動物もある。 |
||
複眼は図形認識能力をもち、 |
複眼は図形認識能力をもち、数多くの'''個眼'''({{lang|la|ommatidium}})というレンズ様の構成単位から[[モザイク画]]のような[[視覚]]を形成する。単眼は主に明暗を感知するなど補助的な機能を担うが、一部の[[クモ]]の単眼は優れた図形認識能力を持ち、例えば[[ハエトリグモ]]の視力は発達しており、内部の[[網膜]]をも動かすことができる<ref name=":33">Rainer F. Foelix (2011). [https://www.academia.edu/13113405/Biology_of_Spiders ''Biology of spiders'' (3rd ed.)]. New York: Oxford University Press. [[ISBN]] [[特別:文献資料/978-0-19-981324-7|978-0-19-981324-7]]. [[Online Computer Library Center|OCLC]] 693776865</ref>。 |
||
眼をもつ節足動物の中で、[[六脚類]]と[[甲殻類]]は基本的に中眼と側眼をあわせもつ。後者の場合、中眼は[[ノープリウス]]幼生期のノープリウス眼に当たる<ref name=":21">{{Cite journal|last=Ortega-Hernández|first=Javier|date=2015-06-15|title=Homology of Head Sclerites in Burgess Shale Euarthropods|url=http://www.sciencedirect.com/science/article/pii/S0960982215004856|journal=Current Biology|volume=25|issue=12|pages=1625-1631|language=en|doi=10.1016/j.cub.2015.04.034|issn=0960-9822}}</ref>。[[多足類]]は全て中眼を欠き、中で[[ゲジ]]類の[[ムカデ]]は側眼が複眼のままで、他のものは複眼が側単眼に変化した。[[鋏角類]]の中で[[ウミグモ]]は中眼のみ、[[カブトガニ類|カブトガニ]]と[[ウミサソリ]]は複眼と中眼の両方、現生の[[クモ綱|クモガタ類]]は複眼を持たず、中眼と複眼由来の側単眼をあわせもち、或いは片方のみを持つ<ref>{{Cite journal|last=Miether|first=Sebastian T.|last2=Dunlop|first2=Jason A.|date=2016/07|title=Lateral eye evolution in the arachnids|url=https://bioone.org/journals/Arachnology/volume-17/issue-2/arac.2006.17.2.103/Lateral-eye-evolution-in-the-arachnids/10.13156/arac.2006.17.2.103.full|journal=Arachnology|volume=17|issue=2|pages=103-119|doi=10.13156/arac.2006.17.2.103|issn=2050-9928}}</ref>。また、[[カンブリア紀]]の化石節足動物をはじめとし、[[三葉虫]]類や基盤的な節足動物と思われる[[ラディオドンタ類]]も発達した複眼を持つ<ref>{{Cite journal|last=Kühl|first=Gabriele|last2=Briggs|first2=Derek E. G.|last3=Rust|first3=Jes|date=2009-02-06|title=A Great-Appendage Arthropod with a Radial Mouth from the Lower Devonian Hunsrück Slate, Germany|url=http://science.sciencemag.org/content/323/5915/771|journal=Science|volume=323|issue=5915|pages=771-773|language=en|doi=10.1126/science.1166586|issn=0036-8075|pmid=19197061}}</ref><ref>{{Cite journal|last=Paterson|first=John R.|last2=García-Bellido|first2=Diego C.|last3=Lee|first3=Michael S. Y.|last4=Brock|first4=Glenn A.|last5=Jago|first5=James B.|last6=Edgecombe|first6=Gregory D.|date=2011-12|title=Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes|url=https://www.nature.com/articles/nature10689|journal=Nature|volume=480|issue=7376|pages=237–240|language=En|doi=10.1038/nature10689|issn=0028-0836}}</ref><ref name=":25">{{Cite journal|last=Cong|first=Peiyun|last2=Ma|first2=Xiaoya|last3=Hou|first3=Xianguang|last4=Edgecombe|first4=Gregory D.|last5=Strausfeld|first5=Nicholas J.|date=2014-07-16|title=Brain structure resolves the segmental affinity of anomalocaridid appendages|url=https://doi.org/10.1038/nature13486|journal=Nature|volume=513|issue=7519|pages=538–542|language=En|doi=10.1038/nature13486|issn=0028-0836}}</ref><ref>{{Cite journal|last=Strausfeld|author=|first=Nicholas J.|last2=Ma|first2=Xiaoya|last3=Edgecombe|first3=Gregory D.|last4=Fortey|first4=Richard A.|last5=Land|first5=Michael F.|last6=Liu|first6=Yu|last7=Cong|first7=Peiyun|last8=Hou|first8=Xianguang|year=|date=2016-03|title=Arthropod eyes: The early Cambrian fossil record and divergent evolution of visual systems|url=https://www.researchgate.net/publication/281055144_Arthropod_Eyes_The_Early_Cambrian_Fossil_Record_and_Divergent_Evolution_of_Visual_Systems|journal=Arthropod Structure & Development|volume=45|issue=2|page=|pages=152–172|language=en|doi=10.1016/j.asd.2015.07.005}}</ref>。 |
眼をもつ節足動物の中で、[[六脚類]]と[[甲殻類]]は基本的に中眼と側眼をあわせもつ。後者の場合、中眼は[[ノープリウス]]幼生期のノープリウス眼に当たる<ref name=":21">{{Cite journal|last=Ortega-Hernández|first=Javier|date=2015-06-15|title=Homology of Head Sclerites in Burgess Shale Euarthropods|url=http://www.sciencedirect.com/science/article/pii/S0960982215004856|journal=Current Biology|volume=25|issue=12|pages=1625-1631|language=en|doi=10.1016/j.cub.2015.04.034|issn=0960-9822}}</ref>。[[多足類]]は全て中眼を欠き、中で[[ゲジ]]類の[[ムカデ]]は側眼が複眼のままで、他のものは複眼が側単眼に変化した。[[鋏角類]]の中で[[ウミグモ]]は中眼のみ、[[カブトガニ類|カブトガニ]]と[[ウミサソリ]]は複眼と中眼の両方、現生の[[クモ綱|クモガタ類]]は複眼を持たず、中眼と複眼由来の側単眼をあわせもち、或いは片方のみを持つ<ref name=":38">{{Cite journal|last=Miether|first=Sebastian T.|last2=Dunlop|first2=Jason A.|date=2016/07|title=Lateral eye evolution in the arachnids|url=https://bioone.org/journals/Arachnology/volume-17/issue-2/arac.2006.17.2.103/Lateral-eye-evolution-in-the-arachnids/10.13156/arac.2006.17.2.103.full|journal=Arachnology|volume=17|issue=2|pages=103-119|doi=10.13156/arac.2006.17.2.103|issn=2050-9928}}</ref>。また、[[カンブリア紀]]の化石節足動物をはじめとし、[[三葉虫]]類や基盤的な節足動物と思われる[[ラディオドンタ類]]も発達した複眼を持つ<ref>{{Cite journal|last=Kühl|first=Gabriele|last2=Briggs|first2=Derek E. G.|last3=Rust|first3=Jes|date=2009-02-06|title=A Great-Appendage Arthropod with a Radial Mouth from the Lower Devonian Hunsrück Slate, Germany|url=http://science.sciencemag.org/content/323/5915/771|journal=Science|volume=323|issue=5915|pages=771-773|language=en|doi=10.1126/science.1166586|issn=0036-8075|pmid=19197061}}</ref><ref>{{Cite journal|last=Paterson|first=John R.|last2=García-Bellido|first2=Diego C.|last3=Lee|first3=Michael S. Y.|last4=Brock|first4=Glenn A.|last5=Jago|first5=James B.|last6=Edgecombe|first6=Gregory D.|date=2011-12|title=Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes|url=https://www.nature.com/articles/nature10689|journal=Nature|volume=480|issue=7376|pages=237–240|language=En|doi=10.1038/nature10689|issn=0028-0836}}</ref><ref name=":25">{{Cite journal|last=Cong|first=Peiyun|last2=Ma|first2=Xiaoya|last3=Hou|first3=Xianguang|last4=Edgecombe|first4=Gregory D.|last5=Strausfeld|first5=Nicholas J.|date=2014-07-16|title=Brain structure resolves the segmental affinity of anomalocaridid appendages|url=https://doi.org/10.1038/nature13486|journal=Nature|volume=513|issue=7519|pages=538–542|language=En|doi=10.1038/nature13486|issn=0028-0836}}</ref><ref>{{Cite journal|last=Strausfeld|author=|first=Nicholas J.|last2=Ma|first2=Xiaoya|last3=Edgecombe|first3=Gregory D.|last4=Fortey|first4=Richard A.|last5=Land|first5=Michael F.|last6=Liu|first6=Yu|last7=Cong|first7=Peiyun|last8=Hou|first8=Xianguang|year=|date=2016-03|title=Arthropod eyes: The early Cambrian fossil record and divergent evolution of visual systems|url=https://www.researchgate.net/publication/281055144_Arthropod_Eyes_The_Early_Cambrian_Fossil_Record_and_Divergent_Evolution_of_Visual_Systems|journal=Arthropod Structure & Development|volume=45|issue=2|page=|pages=152–172|language=en|doi=10.1016/j.asd.2015.07.005}}</ref>。 |
||
<gallery> |
<gallery> |
||
ファイル:Mantis Shrimp Eyes.JPG|[[眼柄]]に付いた[[モンハナシャコ]]の[[複眼]] |
ファイル:Mantis Shrimp Eyes.JPG|[[眼柄]]に付いた[[モンハナシャコ]]の[[複眼]] |
||
| 113行目: | 117行目: | ||
*[[多足類]]は全て陸上性で、[[六脚類]]と似たような気管と気門で呼吸する。 |
*[[多足類]]は全て陸上性で、[[六脚類]]と似たような気管と気門で呼吸する。 |
||
*[[鋏角類]]の中で、水生の[[カブトガニ類]]は後体の蓋板にある[[書鰓]]で呼吸をし、陸生の[[クモ綱|クモガタ類]]は主に[[書肺]]や気管(気門)を通じて行う。なお、[[ウミグモ綱|ウミグモ]]・[[コヨリムシ]]と一部の[[ダニ]]は呼吸器官を持たず、体表で直接的にガス交換を行う。 |
*[[鋏角類]]の中で、水生の[[カブトガニ類]]は後体の蓋板にある[[書鰓]]で呼吸をし、陸生の[[クモ綱|クモガタ類]]は主に[[書肺]]や気管(気門)を通じて行う。なお、[[ウミグモ綱|ウミグモ]]・[[コヨリムシ]]と一部の[[ダニ]]は呼吸器官を持たず、体表で直接的にガス交換を行う。 |
||
*[[甲殻類]]は通常、付属肢における外葉などの附属体が鰓となって水中に呼吸する。陸生の[[ワラジムシ亜目]]は腹肢にある白体(偽気管)で呼吸し、[[ヤシガニ]]の鰓室は陸上での呼吸に用いられる無数の突起物がある<ref>{{Cite journal|last=Farrelly|first=C.A.|last2=Greenaway|first2=P.|date=2005-1|title=The morphology and vasculature of the respiratory organs of terrestrial hermit crabs (Coenobita and Birgus): gills, branchiostegal lungs and abdominal lungs|url=https://linkinghub.elsevier.com/retrieve/pii/S1467803904000684|journal=Arthropod Structure & Development|volume=34|issue=1|pages=63–87|language=en|doi=10.1016/j.asd.2004.11.002}}</ref>。 |
*[[甲殻類]]は通常、付属肢における外葉などの附属体が鰓となって水中に呼吸する<ref name=":37" />。陸生の[[ワラジムシ亜目]]は腹肢にある白体(偽気管)で呼吸し、[[ヤシガニ]]の鰓室は陸上での呼吸に用いられる無数の突起物がある<ref>{{Cite journal|last=Farrelly|first=C.A.|last2=Greenaway|first2=P.|date=2005-1|title=The morphology and vasculature of the respiratory organs of terrestrial hermit crabs (Coenobita and Birgus): gills, branchiostegal lungs and abdominal lungs|url=https://linkinghub.elsevier.com/retrieve/pii/S1467803904000684|journal=Arthropod Structure & Development|volume=34|issue=1|pages=63–87|language=en|doi=10.1016/j.asd.2004.11.002}}</ref>。 |
||
<gallery> |
<gallery> |
||
ファイル:Actias selene 5th instar spiracles sjh.jpg|[[ネッタイオナガミズアオ]]の[[幼虫]]の[[気門]](矢印先) |
ファイル:Actias selene 5th instar spiracles sjh.jpg|[[ネッタイオナガミズアオ]]の[[幼虫]]の[[気門]](矢印先) |
||
| 127行目: | 131行目: | ||
求愛行動・交尾或いは交接・メイトガード・護卵など、節足動物は分類群によって様々な[[繁殖行動]]を持つ。原則として[[有性生殖]]を行う[[卵生]]動物であるが、[[サソリ]]や[[アブラムシ]]などから[[単為生殖]]と[[卵胎生]]の例も知られる。 |
求愛行動・交尾或いは交接・メイトガード・護卵など、節足動物は分類群によって様々な[[繁殖行動]]を持つ。原則として[[有性生殖]]を行う[[卵生]]動物であるが、[[サソリ]]や[[アブラムシ]]などから[[単為生殖]]と[[卵胎生]]の例も知られる。 |
||
{{wide image|Aeshna cyanea freshly slipped L2.jpg|1000px|[[トンボ]]の[[羽化]]}} |
{{wide image|Aeshna cyanea freshly slipped L2.jpg|1000px|[[トンボ]]の[[羽化]]}} |
||
成長に伴い古い[[外骨格]]の下で新しい外骨格を形成し、'''[[脱皮]]'''([[:en:Ecdysis|Ecdysis]])を通じて古い外骨格から抜け出して成長する。新しい外骨格は柔らかく、固くなるのも時間が掛かり、脱皮直後の節足動物は無防備である。そのため節足動物の脱皮は常に隠れ場所で行うことが多い。中でも古い外骨格を摂食して栄養を回収する種類や、一部の[[クモ]]は脱皮直後のメスを狙って交接することが知られる。 |
成長に伴い古い[[外骨格]]の下で新しい外骨格を形成し、'''[[脱皮]]'''([[:en:Ecdysis|Ecdysis]])を通じて古い外骨格から抜け出して成長する。新しい外骨格は柔らかく、固くなるのも時間が掛かり、脱皮直後の節足動物は無防備である。そのため節足動物の脱皮は常に隠れ場所で行うことが多い。中でも古い外骨格を摂食して栄養を回収する種類や、一部の[[クモ]]は脱皮直後のメスを狙って交接することが知られる<ref name=":33">Rainer F. Foelix (2011). [https://www.academia.edu/13113405/Biology_of_Spiders ''Biology of spiders'' (3rd ed.)]. New York: Oxford University Press. [[ISBN]] [[特別:文献資料/978-0-19-981324-7|978-0-19-981324-7]]. [[Online Computer Library Center|OCLC]] 693776865</ref>。 |
||
節足動物の幼生は基本的に成体と似たような外見を持つが、[[甲殻類]]の[[ノープリウス幼生]]や[[昆虫]]の[[幼虫]]など、成長の過程で著しく形態が変化する[[変態]]を行う分類群も少なくない。[[昆虫]]の成虫になる脱皮過程は'''[[羽化]]'''({{lang|en|Eclosion}})と呼ばれる。一部の節足動物、例えば[[多足類]]の中には、成体になるまで脱皮に伴って体節と脚の数を増やせ、いわゆる増節変態を行うものがある。 |
節足動物の幼生は基本的に成体と似たような外見を持つが、[[甲殻類]]の[[ノープリウス幼生]]や[[昆虫]]の[[幼虫]]など、成長の過程で著しく形態が変化する[[変態]](metamorphosis)を行う分類群も少なくない。[[昆虫]]の成虫になる脱皮過程は'''[[羽化]]'''({{lang|en|Eclosion}})と呼ばれる。一部の節足動物、例えば[[多足類]]の中には、成体になるまで脱皮に伴って体節と脚の数を増やせ、いわゆる[[増節変態]](anamorphic development)を行うものがある<ref>{{Cite journal|last=Edgecombe|first=Gregory D.|last2=Giribet|first2=Gonzalo|date=2007-01-01|title=Evolutionary Biology of Centipedes (Myriapoda: Chilopoda)|url=https://www.annualreviews.org/doi/10.1146/annurev.ento.52.110405.091326|journal=Annual Review of Entomology|volume=52|issue=1|pages=151–170|doi=10.1146/annurev.ento.52.110405.091326|issn=0066-4170}}</ref>。 |
||
<gallery> |
<gallery> |
||
File:Multi-Modal-Courtship-in-the-Peacock-Spider-Maratus-volans-(O.P.-Cambridge-1874)-pone.0025390.s001.ogv|[[ハエトリグモ]]の求愛ダンス |
File:Multi-Modal-Courtship-in-the-Peacock-Spider-Maratus-volans-(O.P.-Cambridge-1874)-pone.0025390.s001.ogv|[[ハエトリグモ]]の求愛ダンス |
||
| 141行目: | 145行目: | ||
== 他の動物門との関係性 == |
== 他の動物門との関係性 == |
||
{{see also|脱皮動物|汎節足動物|側節足動物}} |
|||
{{cladogram |
{{cladogram |
||
|title= |
|title= |
||
| 155行目: | 158行目: | ||
|2={{clade |
|2={{clade |
||
|label1=[[冠輪動物]] |
|label1=[[冠輪動物]] |
||
|1=[[File:Annelida collage.png|80px]] [[環形動物門]]、[[File: |
|1=[[File:Annelida collage.png|80px]] [[環形動物門]]、[[File:Mollusk collage.jpg|80px]] [[軟体動物門]]など |
||
|label2=[[脱皮動物]] |
|label2=[[脱皮動物]] |
||
|2={{clade |
|2={{clade |
||
| 165行目: | 168行目: | ||
|2=[[File:Hypsibiusdujardini.jpg|80px]] [[緩歩動物門]] |
|2=[[File:Hypsibiusdujardini.jpg|80px]] [[緩歩動物門]] |
||
|3=[[File:Arthropoda.jpg|80px]] '''節足動物門''' |
|3=[[File:Arthropoda.jpg|80px]] '''節足動物門''' |
||
}} }} }} }} }} }}{{see also|汎節足動物|脱皮動物|側節足動物}}節足動物と他の[[動物]][[門 (分類学)|門]]の類縁関係は長らく議論をなされ、20世紀末と2000年代を介して劇的に更新された<ref name=":39">{{Cite journal|last=Telford|first=Maximilian J|last2=Bourlat|first2=Sarah J|last3=Economou|first3=Andrew|last4=Papillon|first4=Daniel|last5=Rota-Stabelli|first5=Omar|date=2008-04-27|title=The evolution of the Ecdysozoa|url=https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2614232/|journal=Philosophical Transactions of the Royal Society B: Biological Sciences|volume=363|issue=1496|pages=1529–1537|doi=10.1098/rstb.2007.2243|issn=0962-8436|pmid=18192181|pmc=2614232}}</ref><ref name=":40">{{Cite journal|last=Edgecombe|first=Gregory D.|date=2009-06|title=Palaeontological and Molecular Evidence Linking Arthropods, Onychophorans, and other Ecdysozoa|url=https://evolution-outreach.biomedcentral.com/articles/10.1007/s12052-009-0118-3|journal=Evolution: Education and Outreach|volume=2|issue=2|pages=178–190|language=en|doi=10.1007/s12052-009-0118-3|issn=1936-6434}}</ref><ref name="stem" /><ref name=":41">{{Cite journal|last=Giribet|first=Gonzalo|last2=Edgecombe|first2=Gregory D.|date=2017-09-01|title=Current Understanding of Ecdysozoa and its Internal Phylogenetic Relationships|url=https://doi.org/10.1093/icb/icx072|journal=Integrative and Comparative Biology|volume=57|issue=3|pages=455–466|doi=10.1093/icb/icx072|issn=1540-7063}}</ref><ref name=":4" />。21世紀以前では、[[前口動物]]で[[体節制]]を持つなどの共通点から、節足動物と[[環形動物]]は近縁である同時に、[[舌形動物]](シタムシ)・[[有爪動物]](カギムシ)・[[緩歩動物]](クマムシ)という3つの動物門は、両者の中間形態を示唆するという考えが主流であった。これらの動物群は、'''[[体節動物]]'''({{Sname|[[:en:Articulata hypothesis|Articulata]]}}<ref>Haeckel E. 1866. Generelle Morphologie der Organismen. Allgemeine Grundzüge der Organischen formen-wissenschaft, mechanisch begründet durch die von Charles Darwin reformirte descendenztheorie, 2 vols. Berlin: Georg Reimer.</ref>)という[[単系統群]]を構成すると考えられ<ref>Snodgrass, Robert Evans. 1938. "[https://repository.si.edu/handle/10088/23666 Evolution of the annelida onychophora and arthropoda]." ''Smithsonian Miscellaneous Collections''. 97 (6):1–159.</ref><ref>{{Cite book|title=Arthropod phylogeny with special reference to insects|url=http://archive.org/details/arthropodphyloge0000boud|publisher=New York : Wiley|date=1979|isbn=978-0-471-04290-7|others=Internet Archive|first=H. Bruce|last=Boudreaux}}</ref><ref>{{Cite journal|last=Dzik|first=Jerzy|last2=Krumbiegel|first2=Günter|date=1989|title=The oldest ‘onychophoran’Xenusion: a link connecting phyla?|url=https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1502-3931.1989.tb01679.x|journal=Lethaia|volume=22|issue=2|pages=169–181|language=en|doi=10.1111/j.1502-3931.1989.tb01679.x|issn=1502-3931}}</ref><ref>{{Cite journal|last=MONGE-NAJERA|first=JULIAN|date=1995-05-01|title=Phylogeny, biogeography and reproductive trends in the Onychophora|url=https://www.researchgate.net/publication/227637906|journal=Zoological Journal of the Linnean Society|volume=114|issue=1|pages=21–60|doi=10.1111/j.1096-3642.1995.tb00111.x|issn=0024-4082}}</ref>、中でも有爪動物と緩歩動物は節足動物に内包され<ref name=":35" /><ref>{{Cite journal|last=LANKESTER|first=E. RAY|date=1904-03-01|title=Memoirs: The Structure and Classification of the Arthropoda|url=https://doi.org/10.1242/jcs.s2-47.188.523|journal=Journal of Cell Science|volume=s2-47|issue=188|pages=523–582|doi=10.1242/jcs.s2-47.188.523|issn=0021-9533}}</ref>、もしくは舌形動物・有爪動物・緩歩動物がまとめて[[側節足動物]]([[:w::Parathropoda|Parathropoda]]<ref>Vandel A. 1949. Arthropodes: Généralités, composition de l’embranchement. In: Grassé pp, ed. Traité de Zoologie, 6. Paris: Masson, 79–158.</ref>)として区別される経緯すらあった<ref>SALVADOR V. PERIS 1971. [http://bba.bioucm.es/cont/docs/peris/DOCS/PERIS39.pdf Los órdenes de los artrópodos. ''Trab. Cátedra Artrópodos''] (Fac. Biología, Univ. Complutense) 1. 1-121</ref>。 |
|||
}} }} }} }} }} }} |
|||
しかし21世紀以降では、[[分子系統学]]をはじめとする多方面([[遺伝子発現]]・[[解剖学]]・[[発生学]])の進展により、環形動物は他の体節動物に類縁でなく、むしろ[[トロコフォア]]幼生を共有する[[軟体動物]]などと共に単系統群の'''[[冠輪動物]]'''([[:en:Lophotrochozoa|Lophotrochozoa]])に属するものであると判明した。同時に他の体節動物も環形動物より、むしろ[[脱皮]]などの性質を共有する[[線形動物]]などと単系統群になると判明し、'''[[脱皮動物]]'''({{sname||Ecdysozoa}}<ref>{{Cite journal|last=Aguinaldo|first=Anna Marie A.|last2=Turbeville|first2=James M.|last3=Linford|first3=Lawrence S.|last4=Rivera|first4=Maria C.|last5=Garey|first5=James R.|last6=Raff|first6=Rudolf A.|last7=Lake|first7=James A.|date=1997-05|title=Evidence for a clade of nematodes, arthropods and other moulting animals|url=https://doi.org/10.1038/387489a0|journal=Nature|volume=387|issue=6632|pages=489–493|language=En|doi=10.1038/387489a0|issn=0028-0836}}</ref>)として区別されるようになった<ref>{{Cite book|last=Adoutte|first=A|title=The new animal phylogeny: Reliability and implications|url=https://www.researchgate.net/publication/12535812_The_new_animal_phylogeny_Reliability_and_implications|date=2000-05-01|volume=97|last2=Balavoine|first2=Guillaume|last3=Lartillot|first3=N|last4=Lespinet|first4=O|last5=Prud'homme|first5=B|last6=de Rosa|first6=R}}</ref><ref name=":39" /><ref name=":40" /><ref name=":41" /><ref name=":4" />。かつて体節動物の根拠とされてきた環形動物と他の体節動物の体節制も、後に発生学と遺伝子発現の違いにより別起源([[収斂進化]])であることが示唆される<ref>{{Cite journal|last=Seaver|first=Elaine C.|last2=Kaneshige|first2=Lori M.|date=2006-01-01|title=Expression of ‘segmentation’ genes during larval and juvenile development in the polychaetes Capitella sp. I and H. elegans|url=https://www.sciencedirect.com/science/article/pii/S0012160605007530|journal=Developmental Biology|volume=289|issue=1|pages=179–194|language=en|doi=10.1016/j.ydbio.2005.10.025|issn=0012-1606}}</ref><ref>{{Cite journal|last=Mayer|first=Georg|date=2006-03-01|title=Origin and differentiation of nephridia in the Onychophora provide no support for the Articulata|url=https://doi.org/10.1007/s00435-005-0006-5|journal=Zoomorphology|volume=125|issue=1|pages=1–12|language=en|doi=10.1007/s00435-005-0006-5|issn=1432-234X}}</ref><ref>{{Cite journal|last=Chipman|first=Ariel D.|date=2010|title=Parallel evolution of segmentation by co-option of ancestral gene regulatory networks|url=https://onlinelibrary.wiley.com/doi/abs/10.1002/bies.200900130|journal=BioEssays|volume=32|issue=1|pages=60–70|language=en|doi=10.1002/bies.200900130|issn=1521-1878}}</ref>。更に、かつて側節足動物ともされてきた群の中で、緩歩動物と有爪動物は節足動物に内包されない独立の動物門として広く認められる一方、舌形動物は独立した動物門ではなく、[[鰓尾類]]に近縁の[[甲殻類]]、すなわち極端に特化した節足動物の一員だと判明した<ref>{{Cite journal|last=Storch|first=Volker|last2=Jamieson|first2=Barrie G. M.|date=1992-02-01|title=Further spermatological evidence for including the pentastomida (tongue worms) in the Crustacea|url=https://www.sciencedirect.com/science/article/pii/002075199290085Y|journal=International Journal for Parasitology|volume=22|issue=1|pages=95–108|language=en|doi=10.1016/0020-7519(92)90085-Y|issn=0020-7519}}</ref><ref>{{Cite journal|last=Lavrov|first=Dennis V.|last2=Brown|first2=Wesley M.|last3=Boore|first3=Jeffrey L.|date=2004-03-07|title=Phylogenetic position of the Pentastomida and (pan)crustacean relationships|url=https://royalsocietypublishing.org/doi/10.1098/rspb.2003.2631|journal=Proceedings of the Royal Society of London. Series B: Biological Sciences|volume=271|issue=1538|pages=537–544|doi=10.1098/rspb.2003.2631|pmid=15129965|pmc=PMC1691615}}</ref><ref name=":4" />。 |
|||
節足動物と他の動物門の類縁関係については、長く議論されていた。以前は、[[前口動物]]であること、[[体節制]]を持つことなどの共通点から、[[環形動物]]に近縁であると考えられた。そして[[舌形動物]]・[[有爪動物]]・[[緩歩動物]]という3つの動物門は、両者の中間形態を示唆するものと思われ、'''[[側節足動物]]'''({{sname||Parathropoda}})としてまとめられた。こうして、[[環形動物]]・[[側節足動物]]・節足動物という[[体節制]]をもつ3つの動物群は、'''[[体節動物]]'''({{Sname|[[:en:Articulata hypothesis|Articulata]]}})という[[単系統群]]を構成すると考えられた。その中でも[[有爪動物]]と[[緩歩動物]]は、節足動物に内包されたという経緯もあった<ref name="stem" /><ref name="Legg2013" />。 |
|||
| ⚫ | その結果、古典的な「体節動物」と「側節足動物」はいずれも系統関係を反映できない[[多系統群]]として解体され、徐々に21世紀以降の分類体系から廃止された<ref name=":41" /><ref name=":4" />。21世紀、特に2000年代後期以降では、脱皮動物の中で、節足動物・有爪動物・緩歩動物という3動物門が単系統群を構成する説が広く認められ、まとめて'''[[汎節足動物]]'''({{sname||Panarthropoda}}<ref>Nielsen, C. (1995). Animal Evolution, Interrelationships of the Living Phyla. Oxford University Press, Oxford.</ref>)と呼ばれている<ref>{{Cite journal|last=Persson|first=Dennis K.|last2=Halberg|first2=Kenneth A.|last3=Jørgensen|first3=Aslak|last4=Møbjerg|first4=Nadja|last5=Kristensen|first5=Reinhardt M.|date=2012-11-01|title=Neuroanatomy of ''Halobiotus crispae'' (Eutardigrada: Hypsibiidae): Tardigrade brain structure supports the clade panarthropoda|url=http://onlinelibrary.wiley.com/doi/10.1002/jmor.20054/abstract|journal=Journal of Morphology|volume=273|issue=11|language=en|doi=10.1002/jmor.20054/abstract|issn=1097-4687}}</ref><ref>{{Cite journal|last=Hyun Ryu|first=Shi|last2=Lee|first2=Jimin|last3=Jang|first3=Kuem-Hee|last4=Hwa Choi|first4=Eun|last5=Ju Park|first5=Shin|last6=Chang|first6=Cheon|last7=Kim|first7=Won|last8=Hwang|first8=Ui Wook|date=2008-01-01|title=Partial mitochondrial gene arrangements support a close relationship between Tardigrada and Arthropoda|url=https://www.researchgate.net/publication/5667274_Partial_mitochondrial_gene_arrangements_support_a_close_relationship_between_Tardigrada_and_Arthropoda|journal=Molecules and cells|volume=24|pages=351-357}}</ref><ref name="Panarthropoda">{{cite journal |author=Omar Rota-Stabelli, Ehsan Kayal, Dianne Gleeson, Jennifer Daub, Jeffrey L. Boore, Maximilian J. Telford, Davide Pisani, Mark Blaxter & Dennis V. Lavrov |year=2010 |title=Ecdysozoan mitogenomics: evidence for a common origin of the legged invertebrates, the Panarthropoda |journal=Genome Biology and Evolution |volume=2 |pages=425–440 |doi=10.1093/gbe/evq030 |pmid=20624745 |url=http://gbe.oxfordjournals.org/content/2/425}}</ref><ref name=":4" />。 |
||
しかしその後は、[[発生学]]・[[分岐分類学]]・[[分子系統学]]・[[分子遺伝学]]など多方面の情報により、環形動物は節足動物に類縁でなく、むしろ[[トロコフォア]]幼生を共有する[[軟体動物]]などと共に単系統群の'''[[冠輪動物]]'''に属するものであると判明した。したがって体節制という共通点は、単なる前口動物の[[祖先形質]]もしくは[[収斂進化]]の結果と思われ、節足動物と[[環形動物]]の類縁関係は否定的になった<ref>{{Cite journal|last=Aguinaldo|first=Anna Marie A.|last2=Turbeville|first2=James M.|last3=Linford|first3=Lawrence S.|last4=Rivera|first4=Maria C.|last5=Garey|first5=James R.|last6=Raff|first6=Rudolf A.|last7=Lake|first7=James A.|date=1997-05|title=Evidence for a clade of nematodes, arthropods and other moulting animals|url=https://doi.org/10.1038/387489a0|journal=Nature|volume=387|issue=6632|pages=489–493|language=En|doi=10.1038/387489a0|issn=0028-0836}}</ref><ref>{{Cite book|last=Adoutte|first=A|title=The new animal phylogeny: Reliability and implications|url=https://www.researchgate.net/publication/12535812_The_new_animal_phylogeny_Reliability_and_implications|date=2000-05-01|volume=97|last2=Balavoine|first2=Guillaume|last3=Lartillot|first3=N|last4=Lespinet|first4=O|last5=Prud'homme|first5=B|last6=de Rosa|first6=R}}</ref>。[[舌形動物]]は、のちに[[鰓尾類]]に近縁の節足動物であると解明されており、[[緩歩動物]]と[[有爪動物]]も、他の動物門より節足動物に近縁と見なされるようになった。その結果として[[側節足動物]]は[[多系統]]となり、否定された。 |
|||
なお、脱皮動物と汎節足動物の単系統性が広く認められるものの、汎節足動物と他の脱皮動物([[環神経動物]] [[:en:Cycloneuralia|Cycloneuralia]]<ref>{{Cite book|title=Ultrastruktur und Phylogenie von Seison nebaliae (Grube 1859) und Seison annulatus (Claus 1876) : Hypothesen zu phylogenetischen Verwandtschaftsverhältnissen innerhalb der Bilateria|url=https://www.worldcat.org/title/ultrastruktur-und-phylogenie-von-seison-nebaliae-grube-1859-und-seison-annulatus-claus-1876-hypothesen-zu-phylogenetischen-verwandtschaftsverhaltnissen-innerhalb-der-bilateria/oclc/40302795|publisher=Cuvillier|date=1995|isbn=978-3-89588-295-1|oclc=40302795|language=German|first=Wilko|last=Ahlrichs}}</ref><ref>{{Citation|和書|title=Cycloneuralia|url=https://www.researchgate.net/publication/281003133|publisher=Springer|date=2015|accessdate=2021-12-23|isbn=978-3-7091-1865-8|pages=1–13|doi=10.1007/978-3-7091-1865-8_1|language=en|first=Andreas|last=Hejnol|editor-first=Andreas|editor-last=Wanninger|year=}}</ref>)の系統関係ははっきりしておらず、汎節足動物内の3動物門は形態の類似と分子系統解析の食い違いにより、お互いの系統関係は諸説に分かれている<ref name="PanarthroHypothesis">{{Cite journal|date=2017-05-01|title=Segmentation in Tardigrada and diversification of segmental patterns in Panarthropoda|url=https://www.sciencedirect.com/science/article/pii/S1467803916301487|journal=Arthropod Structure & Development|volume=46|issue=3|pages=328–340|language=en|doi=10.1016/j.asd.2016.10.005|issn=1467-8039}}</ref>。これらの議論の詳細については[[汎節足動物#系統関係]]および[[汎節足動物#内部系統関係]]を参照のこと。<gallery> |
|||
| ⚫ | 21世紀 |
||
| ⚫ | |||
ファイル:CelegansGoldsteinLabUNC.jpg|[[センチュウ]]([[線形動物]])は汎節足動物と同様に[[脱皮]]で成長する。 |
|||
なお、有爪動物と緩歩動物にはそれぞれ節足動物と共通の別の形質が見られることをはじめとして汎節足動物の内部の系統関係は諸説に分かれる<ref name="PanarthroHypothesis">{{Cite journal|date=2017-05-01|title=Segmentation in Tardigrada and diversification of segmental patterns in Panarthropoda|url=https://www.sciencedirect.com/science/article/pii/S1467803916301487|journal=Arthropod Structure & Development|volume=46|issue=3|pages=328–340|language=en|doi=10.1016/j.asd.2016.10.005|issn=1467-8039}}</ref>。この議論の詳細については[[汎節足動物#内部系統]]を参照のこと。 |
|||
ファイル:Armillifer armillatus.jpg|[[シタムシ]]の外見は節足動物らしからぬ、かつて独自の動物門([[舌形動物]])とされていた。 |
|||
<gallery> |
|||
| ⚫ | |||
| ⚫ | |||
ファイル:Tardigrade zenuw.jpg|[[クマムシ]]([[緩歩動物]])の胴部は節足動物に似た神経節をもつ。 |
|||
| ⚫ | |||
File:Hypsibiusdujardini.jpg|[[クマムシ]]([[緩歩動物]])の胴部は節足動物らしい神経節をもつ。 |
|||
</gallery> |
</gallery> |
||
== 起源 == |
== 起源 == |
||
{{ |
{{See also|ラディオドンタ類#系統関係|Dinocaridida#系統関係|葉足動物#節足動物との関係性|汎節足動物#起源と進化}}{{cladogram |
||
{{cladogram |
|||
|title= |
|title= |
||
|align=right |
|align=right |
||
| 208行目: | 208行目: | ||
|3=[[鋏角類]] [[ファイル:Chelicerata Collage.png|80px]] |
|3=[[鋏角類]] [[ファイル:Chelicerata Collage.png|80px]] |
||
|4=†[[:en:Artiopoda|Artiopoda]] [[ファイル:Olenoides serratus 3d.jpg|80px]] |
|4=†[[:en:Artiopoda|Artiopoda]] [[ファイル:Olenoides serratus 3d.jpg|80px]] |
||
|5=†[[フーシェンフイア類]] [[ |
|5=†[[フーシェンフイア類]] [[ファイル:20211117 Fuxianhuiida Fuxianhuiids.png|80px]] |
||
|6=†[[:en:Hymenocarina|Hymenocarina]] [[ファイル:20211025 Waptia fieldensis.png|80px]] |
|6=†[[:en:Hymenocarina|Hymenocarina]] [[ファイル:20211025 Waptia fieldensis.png|80px]] |
||
|7=[[多足類]]、[[甲殻類]]、[[六脚類]] [[ファイル:Myriapod collage.png|70px]][[ファイル:Crustacea.jpg|70px]][[ファイル:Insect collage.png|70px]] |
|7=[[多足類]]、[[甲殻類]]、[[六脚類]] [[ファイル:Myriapod collage.png|70px]][[ファイル:Crustacea.jpg|70px]][[ファイル:Insect collage.png|70px]] |
||
}} }} }} }} }} }} }} }} |
}} }} }} }} }} }} }} }} |
||
知られる最古の節足動物は、およそ5億年前の[[カンブリア紀]]に遡る<ref name="Daley2018" />。この[[地質時代]]からは、[[アノマロカリス]]をはじめとする[[ラディオドンタ類]]([[アノマロカリス類]])や[[オパビニア]]など、節足動物的な形質と節足動物らしからぬ形質を掛け合わせた[[古生物]]が見つかり、節足動物の初期系統を表したものとして広く注目される(後述)<ref name=":23">{{Cite journal|last=Budd|first=Graham|date=1993-08|title=A Cambrian gilled lobopod from Greenland|url=https://doi.org/10.1038/364709a0|journal=Nature|volume=364|issue=6439|pages=709–711|language=En|doi=10.1038/364709a0|issn=0028-0836}}</ref><ref>{{Cite journal|last=BUDD|first=GRAHAM E.|date=1996-03|title=The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group|url=https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1502-3931.1996.tb01831.x|journal=Lethaia|volume=29|issue=1|pages=1–14|language=en|doi=10.1111/j.1502-3931.1996.tb01831.x|issn=0024-1164}}</ref><ref name=":25" /><ref name=":6">{{Cite journal|last=Van Roy|first=Peter|last2=Daley|first2=Allison C.|last3=Briggs|first3=Derek E. G.|date=2015-03-11|title=Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps|url=https://www.nature.com/articles/nature14256|journal=Nature|volume=522|issue=7554|pages=77–80|language=En|doi=10.1038/nature14256|issn=0028-0836}}</ref><ref name=":4" /><ref name=":26">{{Cite journal|last=Edgecombe|first=Gregory D.|date=2020-11-02|title=Arthropod Origins: Integrating Paleontological and Molecular Evidence|url=https://www.annualreviews.org/doi/10.1146/annurev-ecolsys-011720-124437|journal=Annual Review of Ecology, Evolution, and Systematics|volume=51|issue=1|pages=1–25|doi=10.1146/annurev-ecolsys-011720-124437|issn=1543-592X}}</ref>。しかし、派生的な節足動物の最古の[[化石]]記録はそれ以上に早期(体の化石は約5億2,100万年前まで、[[生痕化石]]は約5億3,700万年前まで) |
知られる最古の節足動物は、およそ5億年前の[[カンブリア紀]]に遡る<ref name="Daley2018" />。この[[地質時代]]からは、[[アノマロカリス]]をはじめとする[[ラディオドンタ類]]([[アノマロカリス類]])や[[オパビニア]]など、節足動物的な形質と節足動物らしからぬ形質を掛け合わせた[[古生物]]が見つかり、節足動物の初期系統を表したものとして広く注目される(後述)<ref name=":23">{{Cite journal|last=Budd|first=Graham|date=1993-08|title=A Cambrian gilled lobopod from Greenland|url=https://doi.org/10.1038/364709a0|journal=Nature|volume=364|issue=6439|pages=709–711|language=En|doi=10.1038/364709a0|issn=0028-0836}}</ref><ref>{{Cite journal|last=BUDD|first=GRAHAM E.|date=1996-03|title=The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group|url=https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1502-3931.1996.tb01831.x|journal=Lethaia|volume=29|issue=1|pages=1–14|language=en|doi=10.1111/j.1502-3931.1996.tb01831.x|issn=0024-1164}}</ref><ref name=":25" /><ref name=":6">{{Cite journal|last=Van Roy|first=Peter|last2=Daley|first2=Allison C.|last3=Briggs|first3=Derek E. G.|date=2015-03-11|title=Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps|url=https://www.nature.com/articles/nature14256|journal=Nature|volume=522|issue=7554|pages=77–80|language=En|doi=10.1038/nature14256|issn=0028-0836}}</ref><ref name=":4" /><ref name=":26">{{Cite journal|last=Edgecombe|first=Gregory D.|date=2020-11-02|title=Arthropod Origins: Integrating Paleontological and Molecular Evidence|url=https://www.annualreviews.org/doi/10.1146/annurev-ecolsys-011720-124437|journal=Annual Review of Ecology, Evolution, and Systematics|volume=51|issue=1|pages=1–25|doi=10.1146/annurev-ecolsys-011720-124437|issn=1543-592X}}</ref>。しかし、派生的な節足動物の最古の[[化石]]記録はそれ以上に早期(体の化石は約5億2,100万年前まで、[[生痕化石]]は約5億3,700万年前まで)のため、節足動物の初期系統は中間化石記録を欠いており、実際の起源は記録以上に古かったことが示唆される<ref name="Daley2018">{{Cite journal|last=Daley|first=Allison|last2=Antcliffe|first2=Jonathan|last3=Drage|first3=Harriet|last4=Pates|first4=Stephen|date=2018-05-21|title=Early fossil record of Euarthropoda and the Cambrian Explosion|url=https://www.researchgate.net/publication/325275404_Early_fossil_record_of_Euarthropoda_and_the_Cambrian_Explosion?ev=publicSearchHeader&_sg=w2vqyEGTVn0XGu-foabSSqxq-ic8EqYvx-1JQtfkTbNjup2t2X7h5LVXhaRoRN1dHbsiov_gjm2WrY8|journal=Proceedings of the National Academy of Sciences|volume=115|pages=201719962|doi=10.1073/pnas.1719962115}}</ref>。なお、カンブリア紀とその直前の[[エディアカラ紀]]の[[ラーガーシュテッテ|化石産地]]の比較により、その起源は、エディアカラ紀晩期(約5億5,000万年前)より古くはなかったと考えられる<ref name="Daley2018" />。 |
||
<gallery mode="packed" heights="120"> |
<gallery mode="packed" heights="120"> |
||
| 229行目: | 229行目: | ||
ファイル:20210310 Kylinxia zhangi.png|[[ラディオドンタ類]]と[[オパビニア]]に似た頭部構造をもつ真節足動物[[キリンシア]] |
ファイル:20210310 Kylinxia zhangi.png|[[ラディオドンタ類]]と[[オパビニア]]に似た頭部構造をもつ真節足動物[[キリンシア]] |
||
ファイル:Isoxys auritus.jpg|[[イソキシス類]]の[[イソキシス]] |
ファイル:Isoxys auritus.jpg|[[イソキシス類]]の[[イソキシス]] |
||
ファイル:Guangweicaris restoration.png|[[フーシェンフイア類]]の[[グ |
ファイル:Guangweicaris restoration.png|[[フーシェンフイア類]]の[[グァンウェイカリス]] |
||
</gallery> |
</gallery> |
||
| 237行目: | 237行目: | ||
=== 系統関係と体節の相同性 === |
=== 系統関係と体節の相同性 === |
||
{{see also|鋏角類#体節と付属肢の対応関係|鋏角類#系統関係|大顎類|多足類#系統関係|汎甲殻類|甲殻類#系統関係|六脚類#分類}} |
{{see also|鋏角類#体節と付属肢の対応関係|鋏角類#系統関係|大顎類|多足類#系統関係|汎甲殻類|甲殻類#系統関係|六脚類#分類}} |
||
現生の節足動物は、'''[[鋏角亜門]]'''([[クモ |
現生の節足動物は、'''[[鋏角亜門]]'''([[クモ]]、[[サソリ]]、[[カブトガニ類|カブトガニ]]など)・'''[[多足亜門]]'''([[ムカデ]]、[[ヤスデ]]など)・'''[[甲殻亜門]]'''([[十脚目|カニとエビ]]、[[フジツボ]]、[[ミジンコ目|ミジンコ]]など)・'''[[六脚亜門]]'''([[昆虫]]、[[トビムシ目|トビムシ]]など)という4つの[[亜門]]に分類されている。[[化石]]種まで範囲を広げれば、[[三葉虫]]などを含んだ'''{{sname||Artiopoda}}'''亜門という過去の大グループや、確定的に現生の亜門に収まらない分類群も数多く知られている([[節足動物#絶滅した分類群|後述参照]])<ref name=":4" /><ref name=":26" />。 |
||
特に注目される |
それそれの分類群で特に注目される特徴は、先頭複数体節の融合でできた頭部(真鋏角類の場合は前体)の[[関節肢|付属肢]]である。その基本構成は各亜門によって異なり、次の通りである: |
||
*[[六脚類]]:[[触角]]1対・[[大顎]]1対・[[小顎]]1対・[[小顎#六脚類|下唇]]1枚(1対の小顎から癒合した部分) |
*[[六脚類]]:[[触角]]1対・[[大顎]]1対・[[小顎]]1対・[[小顎#六脚類|下唇]]1枚(1対の小顎から癒合した部分) |
||
*[[多足類]]:触角1対・大顎1対・小顎2対 |
*[[多足類]]:触角1対・大顎1対・小顎2対 |
||
| 247行目: | 247行目: | ||
*{{sname||Artiopoda}}類:触角1対・脚3対以上 |
*{{sname||Artiopoda}}類:触角1対・脚3対以上 |
||
これらの付属肢の対応関係([[相同|相同性]])は、節足動物の内部系統や他の[[汎節足動物]]との関係性を大きく左右する指標とされ、そこから繰り広げた議論は「[[:en:Arthropod head problem|'''Arthropod head problem''']]」として知られている<ref name="head" />。 |
これらの付属肢の対応関係([[相同|相同性]])は、節足動物の内部系統や他の[[汎節足動物]]との関係性を大きく左右する指標とされ、そこから繰り広げた議論は「[[:en:Arthropod head problem|'''Arthropod head problem''']]」として知られている<ref name="head" /><ref name=":4" />。 |
||
[[ファイル:Chelicerata Collage.png|250px|サムネイル|[[鋏角類]]の体節と付属肢の構成は他の節足動物とは大きく異なる。]] |
[[ファイル:Chelicerata Collage.png|250px|サムネイル|[[鋏角類]]の体節と付属肢の構成は他の節足動物とは大きく異なる。]] |
||
| 255行目: | 255行目: | ||
</gallery> |
</gallery> |
||
古典的な |
21世紀以前の古典的な分類学では、[[鋏角類]]の[[触角]]を欠くという特徴は二次的退化と考えられ<ref name=":27">{{Cite book|title=On the relationships and phylogeny of fossil and recent Arachnomorpha: a comparative study on Arachnida, Xiphosura, Eurypterida, Trilobita, and other fossil Arthropoda|url=https://paleoarchive.com/literature/Stormer1944-RelationshipPhylogenyArachnomorpha.pdf|publisher=Jacob Dybwad|date=1944|location=Oslo|oclc=961296639|language=English|first=Leif|last=Størmer}}</ref>、[[多足類]]と[[六脚類]]は近縁とされてきた<ref>{{Cite book|title=Arthropod Relationships|url=https://doi.org/10.1007/978-94-011-4904-4_22|publisher=Springer Netherlands|date=1998|location=Dordrecht|isbn=978-94-011-4904-4|pages=295–303|doi=10.1007/978-94-011-4904-4_22|language=en|first=O.|last=Kraus|editor-first=R. A.|editor-last=Fortey|editor2-first=R. H.|editor2-last=Thomas}}</ref>。しかしこれらの知見は、後に[[発生学]]・[[神経解剖学]]・[[遺伝子発現]]・[[分子系統学]]など多方面の情報に否定され、代わりに[[鋏角類]]の[[鋏角]]は[[大顎類]]の第1触角に相同で<ref>{{Cite journal|last=Telford|first=Maximilian J.|last2=Thomas|first2=Richard H.|date=1998-09-01|title=Expression of homeobox genes shows chelicerate arthropods retain their deutocerebral segment|url=http://www.pnas.org/content/95/18/10671|journal=Proceedings of the National Academy of Sciences|volume=95|issue=18|pages=10671–10675|language=en|issn=0027-8424|pmid=9724762}}</ref><ref>{{Cite journal|last=Mittmann|first=Beate|last2=Scholtz|first2=Gerhard|date=2003-02-01|title=Development of the nervous system in the "head" of Limulus polyphemus (Chelicerata: Xiphosura): morphological evidence for a correspondence between the segments of the chelicerae and of the (first) antennae of Mandibulata|url=https://doi.org/10.1007/s00427-002-0285-5|journal=Development Genes and Evolution|volume=213|issue=1|pages=9?17|language=en|doi=10.1007/s00427-002-0285-5|issn=1432-041X}}</ref><ref>{{Cite journal|date=2005-07-01|title=Immunohistochemical localization of neurotransmitters in the nervous system of larval Limulus polyphemus (Chelicerata, Xiphosura): evidence for a conserved protocerebral architecture in Euarthropoda|url=https://www.sciencedirect.com/science/article/abs/pii/S1467803905000186|journal=Arthropod Structure & Development|volume=34|issue=3|pages=327–342|language=en|doi=10.1016/j.asd.2005.01.006|issn=1467-8039}}</ref><ref>{{Cite journal|last=Sharma|first=Prashant P.|last2=Tarazona|first2=Oscar A.|last3=Lopez|first3=Davys H.|last4=Schwager|first4=Evelyn E.|last5=Cohn|first5=Martin J.|last6=Wheeler|first6=Ward C.|last7=Extavour|first7=Cassandra G.|date=2015-06-07|title=A conserved genetic mechanism specifies deutocerebral appendage identity in insects and arachnids|url=https://royalsocietypublishing.org/doi/full/10.1098/rspb.2015.0698|journal=Proceedings of the Royal Society B: Biological Sciences|volume=282|issue=1808|pages=20150698|doi=10.1098/rspb.2015.0698|pmid=25948691|pmc=PMC4455815}}</ref>、多足類と六脚類の直接の関連も支持されず、むしろ六脚類は[[側系統群]]の[[甲殻類]]から分岐したことが強く示唆される([[汎甲殻類]]仮説)<ref>{{Cite journal|last=Giribet|first=Gonzalo|last2=Ribera|first2=Carles|date=2000-06|title=A Review of Arthropod Phylogeny: New Data Based on Ribosomal DNA Sequences and Direct Character Optimization|url=https://doi.org/10.1111/j.1096-0031.2000.tb00353.x|journal=Cladistics|volume=16|issue=2|pages=204–231|language=en|doi=10.1111/j.1096-0031.2000.tb00353.x|issn=0748-3007}}</ref><ref>{{Cite journal|last=Nardi|first=Francesco|last2=Spinsanti|first2=Giacomo|last3=Boore|first3=Jeffrey L.|last4=Carapelli|first4=Antonio|last5=Dallai|first5=Romano|last6=Frati|first6=Francesco|date=2003-03-21|title=Hexapod Origins: Monophyletic or Paraphyletic?|url=http://science.sciencemag.org/content/299/5614/1887|journal=Science|volume=299|issue=5614|pages=1887–1889|language=en|doi=10.1126/science.1078607|issn=0036-8075|pmid=12649480}}</ref><ref>{{Cite journal|last=Regier|first=Jerome C.|last2=Shultz|first2=Jeffrey W.|last3=Kambic|first3=Robert E.|date=2005-02-22|title=Pancrustacean phylogeny: hexapods are terrestrial crustaceans and maxillopods are not monophyletic|url=https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1634985/|journal=Proceedings of the Royal Society B: Biological Sciences|volume=272|issue=1561|pages=395–401|doi=10.1098/rspb.2004.2917|issn=0962-8452|pmid=15734694|pmc=1634985}}</ref><ref name=":0">{{Cite journal|last=Regier|first=Jerome C.|last2=Shultz|first2=Jeffrey W.|last3=Zwick|first3=Andreas|last4=Hussey|first4=April|last5=Ball|first5=Bernard|last6=Wetzer|first6=Regina|last7=Martin|first7=Joel W.|last8=Cunningham|first8=Clifford W.|date=2010-02|title=Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences|url=https://doi.org/10.1038/nature08742|journal=Nature|volume=463|issue=7284|pages=1079–1083|language=En|doi=10.1038/nature08742|issn=0028-0836}}</ref><ref name=Oakley>{{Cite journal|last=Oakley|first=Todd H.|last2=Wolfe|first2=Joanna M.|last3=Lindgren|first3=Annie R.|last4=Zaharoff|first4=Alexander K.|date=2012-09-12|title=Phylotranscriptomics to Bring the Understudied into the Fold: Monophyletic Ostracoda, Fossil Placement, and Pancrustacean Phylogeny|url=https://doi.org/10.1093/molbev/mss216|journal=Molecular Biology and Evolution|volume=30|issue=1|pages=215–233|language=en|doi=10.1093/molbev/mss216|issn=1537-1719}}</ref><ref>{{Cite web|url=https://www.semanticscholar.org/paper/Linking-Insects-with-Crustacea-:-Comparative-of-the-Harrison-Sagi/738865517bf8570f77cb89a302ba2c1344f4e00f|title=Linking Insects with Crustacea : Comparative Physiology of the Pancrustacea Organized by|accessdate=2018-10-24|last=Harrison|first=Jon Fewell|date=2015|language=en}}</ref><ref>{{Cite journal|last=Lozano-Fernandez|first=Jesus|last2=Giacomelli|first2=Mattia|last3=Fleming|first3=James F|last4=Chen|first4=Albert|last5=Vinther|first5=Jakob|last6=Thomsen|first6=Philip Francis|last7=Glenner|first7=Henrik|last8=Palero|first8=Ferran|last9=Legg|first9=David A|date=2019-08-01|title=Pancrustacean Evolution Illuminated by Taxon-Rich Genomic-Scale Data Sets with an Expanded Remipede Sampling|url=https://doi.org/10.1093/gbe/evz097|journal=Genome Biology and Evolution|volume=11|issue=8|pages=2055–2070|doi=10.1093/gbe/evz097|issn=1759-6653}}</ref>。また、通常では鋏角類とされる[[ウミグモ]]類の系統位置がしばしば疑問視されるようになり、分子系統解析では鋏角類であることを支持するものが多い<ref>{{Cite book|title=16 Phylogeny of the Chelicerates: Morphological and molecular evidence|url=https://www.degruyter.com/document/doi/10.1515/9783110277524.399/html|publisher=De Gruyter|date=2014-02-27|isbn=978-3-11-027752-4|doi=10.1515/9783110277524.399|language=en|first=Jason|last=Dunlop|first2=Janus|last2=Borner|first3=Thorsten|last3=Burmester}}</ref><ref>{{Cite journal|last=Hassanin|first=Alexandre|last2=Prieur|first2=Stéphanie le|last3=Bonillo|first3=Céline|last4=Krapp|first4=Franz|last5=Corbari|first5=Laure|last6=Sabroux|first6=Romain|date=2017-02-24|title=Biodiversity and phylogeny of Ammotheidae (Arthropoda: Pycnogonida)|url=https://europeanjournaloftaxonomy.eu/index.php/ejt/article/view/410|journal=European Journal of Taxonomy|volume=0|issue=286|language=en|doi=10.5852/ejt.2017.286|issn=2118-9773}}</ref><ref name=":26" />が、2000年代ではそれ以外の節足動物と対置すべき説もある<ref>{{Cite journal|last=Giribet|first=Gonzalo|last2=Edgecombe|first2=Gregory D.|last3=Wheeler|first3=Ward C.|date=2001-09|title=Arthropod phylogeny based on eight molecular loci and morphology|url=https://www.nature.com/articles/35093097|journal=Nature|volume=413|issue=6852|pages=157–161|language=en|doi=10.1038/35093097|issn=1476-4687}}</ref><ref name=":0" /><ref name=Dunlop>{{cite journal |doi=10.1111/j.1439-0469.2004.00284.x |title=Pycnogonid affinities: A review |journal=Journal of Zoological Systematics and Evolutionary Research |volume=43 |pages=8–21 |year=2005 |last1=Dunlop |first1=J. A. |last2=Arango |first2=C. P. }}</ref>。 |
||
[[三葉虫]]類をはじめとして、幾つかの絶滅した節足動物の分類群と現生分類群の類縁関係については、未だに定説がない<ref name=":4" />。例えば[[三葉虫]]などを含む{{sname||Artiopoda}}類は、[[鋏角類]]に類縁という伝統的な系統仮説はあった<ref name=":27" />が、頭部の構成に基づいて触角をもつ現生群(多足類・甲殻類・六脚類)に近い<ref>{{Cite journal|last=Lamsdell|first=James|date=2013-01-01|title=Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura|url=https://www.researchgate.net/publication/259572939_Revised_systematics_of_Palaeozoic_'horseshoe_crabs'_and_the_myth_of_monophyletic_Xiphosura|journal=Zoological Journal of the Linnean Society|volume=167|pages=1–27|doi=10.1111/j.1096-3642.2012.00874.x}}</ref><ref>{{Cite web|url=https://www.researchgate.net/publication/258342467_Heads_Hox_and_the_phylogenetic_position_of_trilobites_Crustacea_and_Arthropod_Phylogeny_-|title=(PDF) Heads, Hox and the phylogenetic position of trilobites. Crustacea and Arthropod Phylogeny. –|accessdate=2018-10-24|website=ResearchGate|language=en}}</ref><ref name=":7">{{Cite journal|last=Scholtz|first=Gerhard|last2=Staude|first2=Andreas|last3=Dunlop|first3=Jason A.|date=2019-06-07|title=Trilobite compound eyes with crystalline cones and rhabdoms show mandibulate affinities|url=https://www.nature.com/articles/s41467-019-10459-8|journal=Nature Communications|volume=10|issue=1|pages=1-7|language=en|doi=10.1038/s41467-019-10459-8|issn=2041-1723}}</ref>、またはどの現生群よりも基盤的<ref name=":8">{{Cite journal|last=Aria|first=Cédric|last2=Caron|first2=Jean-Bernard|date=2017-05|title=Burgess Shale fossils illustrate the origin of the mandibulate body plan|url=https://www.nature.com/articles/nature22080|journal=Nature|volume=545|issue=7652|pages=89–92|language=en|doi=10.1038/nature22080|issn=1476-4687}}</ref><ref name=":4" />とも考えられる。[[メガケイラ類]]に関しては、付属肢と脳の構造に基づいて[[鋏角類]]に近い<ref name=":9">{{Cite journal|last=Tanaka|first=Gengo|last2=Hou|first2=Xianguang|last3=Ma|first3=Xiaoya|last4=Edgecombe|first4=Gregory D.|last5=Strausfeld|first5=Nicholas J.|date=2013-10|title=Chelicerate neural ground pattern in a Cambrian great appendage arthropod|url=https://www.nature.com/articles/nature12520|journal=Nature|volume=502|issue=7471|pages=364–367|language=En|doi=10.1038/nature12520|issn=0028-0836}}</ref>、もしくはどの現生群よりも基盤的<ref name="head" /> |
[[三葉虫]]類をはじめとして、幾つかの絶滅した節足動物の分類群と現生分類群の類縁関係については、未だに定説がない<ref name=":4" />。例えば[[三葉虫]]などを含む{{sname||Artiopoda}}類は、[[鋏角類]]に類縁という伝統的な系統仮説はあった<ref name=":27" />が、頭部の構成に基づいて触角をもつ現生群(多足類・甲殻類・六脚類)に近い<ref>{{Cite journal|last=Lamsdell|first=James|date=2013-01-01|title=Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura|url=https://www.researchgate.net/publication/259572939_Revised_systematics_of_Palaeozoic_'horseshoe_crabs'_and_the_myth_of_monophyletic_Xiphosura|journal=Zoological Journal of the Linnean Society|volume=167|pages=1–27|doi=10.1111/j.1096-3642.2012.00874.x}}</ref><ref>{{Cite web|url=https://www.researchgate.net/publication/258342467_Heads_Hox_and_the_phylogenetic_position_of_trilobites_Crustacea_and_Arthropod_Phylogeny_-|title=(PDF) Heads, Hox and the phylogenetic position of trilobites. Crustacea and Arthropod Phylogeny. –|accessdate=2018-10-24|website=ResearchGate|language=en}}</ref><ref name=":7">{{Cite journal|last=Scholtz|first=Gerhard|last2=Staude|first2=Andreas|last3=Dunlop|first3=Jason A.|date=2019-06-07|title=Trilobite compound eyes with crystalline cones and rhabdoms show mandibulate affinities|url=https://www.nature.com/articles/s41467-019-10459-8|journal=Nature Communications|volume=10|issue=1|pages=1-7|language=en|doi=10.1038/s41467-019-10459-8|issn=2041-1723}}</ref>、またはどの現生群よりも基盤的<ref name=":8">{{Cite journal|last=Aria|first=Cédric|last2=Caron|first2=Jean-Bernard|date=2017-05|title=Burgess Shale fossils illustrate the origin of the mandibulate body plan|url=https://www.nature.com/articles/nature22080|journal=Nature|volume=545|issue=7652|pages=89–92|language=en|doi=10.1038/nature22080|issn=1476-4687}}</ref><ref name=":4" />とも考えられる。[[メガケイラ類]]に関しては、付属肢と脳の構造に基づいて[[鋏角類]]に近い<ref name=":9">{{Cite journal|last=Tanaka|first=Gengo|last2=Hou|first2=Xianguang|last3=Ma|first3=Xiaoya|last4=Edgecombe|first4=Gregory D.|last5=Strausfeld|first5=Nicholas J.|date=2013-10|title=Chelicerate neural ground pattern in a Cambrian great appendage arthropod|url=https://www.nature.com/articles/nature12520|journal=Nature|volume=502|issue=7471|pages=364–367|language=En|doi=10.1038/nature12520|issn=0028-0836}}</ref>、もしくはどの現生群よりも基盤的との説がある<ref name="stem" /><ref name="head" /><ref name=":4" />。 |
||
こうした研究の発展に伴い、節足動物の高次系統に対して様々な系統仮説が与えられており、以下の例が挙げられる<ref name="Legg2013" >{{Cite journal|last=Legg|first=David|date=2013-06|title=The impact of fossils on arthropod phylogeny|url=http://spiral.imperial.ac.uk/handle/10044/1/24168}}</ref>。 |
こうした研究の発展に伴い、節足動物の高次系統に対して様々な系統仮説が与えられており、以下の例が挙げられる<ref name="Legg2013" >{{Cite journal|last=Legg|first=David|date=2013-06|title=The impact of fossils on arthropod phylogeny|url=http://spiral.imperial.ac.uk/handle/10044/1/24168}}</ref><ref name="miyazaki">宮崎勝己「節足動物全体の分類体系・系統の現状」石川良輔編『節足動物の多様性と系統』〈バイオディバーシティ・シリーズ〉第6巻、岩槻邦男・馬渡峻輔監修、裳華房、2008年、11-27頁。</ref>。 |
||
; [[幹性類]] {{sname||Cormogonida}} |
|||
: |
: [[ウミグモ]]類以外の節足動物、すなわち[[真鋏角類]](=ウミグモ類以外の[[鋏角類]])・[[多足類]]・[[甲殻類]]・[[六脚類]]からなる。 |
||
; [[大顎類]] {{sname||Mandibulata}} |
|||
: |
: 多足類・甲殻類・六脚類からなる。大顎を持つことが共有形質とされる。 |
||
; [[汎甲殻類]] {{sname||Pancrustacea}}{{nobold|1=(=[[八分錘類]] {{sname|Tetraconata}})}} |
|||
:甲殻類と六脚類からなる。複眼の八分割される硝子体などが共有形質とされる。 |
|||
; [[多足鋏角類]] {{sname||Myriochelata}}{{nobold|1=(=[[矛盾足類]] {{sname|Paradoxopoda}})}} |
|||
:多足類と鋏角類からなる。 |
|||
; [[裂肢類]] {{sname||Schizoramia}}{{nobold|1=(='CCT' clade)}} |
|||
| ⚫ | :甲殻類・鋏角類・{{sname|Artiopoda}}類などからなる。ニ叉型付属肢が共有形質とされる<ref name=":10">{{Cite book|author=Hou, Xianguang|author2=Bergström, Jan|title=Arthropods of the Lower Cambrian Chengjiang fauna, southwest China|url=https://foreninger.uio.no/ngf/FOS/pdfs/F&S_45.pdf|publisher=Scandinavian University Press|year=1997|location=Oslo|isbn=82-00-37693-1|oclc=38305908}}</ref>。 |
||
| ⚫ | |||
| ⚫ | |||
:多足類と六脚類からなる。気門や頭部の付属肢構成などが共有形質とされる。 |
|||
; {{nobold|広義の}}[[単肢類]] {{sname||Uniramia}} {{nobold|1=<small>''{{lang|la|sensu lato}}''</small>}} |
|||
: |
:[[有爪動物]]・多足類・六脚類からなる。単枝型付属肢が共有形質とされる。またこの系統仮説に従えば、有爪動物は節足動物に含まれ、もしくは節足動物が[[多系統群]]になる。 |
||
; {{sname||Arachnomorpha}}{{nobold|1=(=[[蛛形様類]] {{sname|Arachnata}}<ref name="suzuki">鈴木雄太郎「三葉虫綱」石川良輔編『節足動物の多様性と系統』〈バイオディバーシティ・シリーズ〉第6巻、岩槻邦男・馬渡峻輔監修、裳華房、2008年、114-121頁。</ref>)}} |
|||
| ⚫ | |||
: {{sname||Artiopoda}}類と鋏角類からなる。 |
|||
::[[有爪動物]]・多足類・六脚類からなる。単枝型付属肢が共有形質とされる。またこの系統仮説に従えば、有爪動物は節足動物に含まれ、もしくは節足動物が[[多系統群]]になる。 |
|||
; {{sname||Antennulata}} |
|||
: |
: {{sname|Artiopoda}}類・多足類・甲殻類・六脚類からなる。中大脳性(第1体節由来)の第1触角を持つことが共有形質とされる。 |
||
; [[板肢類]] {{sname||Lamellipedia}}<ref name=":10" /><ref name="suzuki" /> |
|||
* '''{{sname||Antennulata}}''' |
|||
: |
: {{sname|Artiopoda}}類と{{sname|Marrellomorpha}}類からなる。 |
||
2010年代以降では、少なくとも節足動物は有爪動物を含まない単系統群であることや、大顎類説、汎甲殻類説、およびウミグモ類が鋏角類に含まれる説が広く認められる<ref name=":4" />。 |
2010年代以降では、少なくとも節足動物は有爪動物を含まない単系統群であることや、大顎類説、汎甲殻類説、およびウミグモ類が鋏角類に含まれる説が広く認められる<ref name=":4" /><ref name=":26" />。 |
||
{{cladogram |
{{cladogram |
||
|title=節足動物の内部系統関係 |
|title=節足動物の内部系統関係 |
||
| 300行目: | 302行目: | ||
|label1=[[ウミグモ綱]]|1=[[File:Callipallene brevirostris (YPM IZ 077244) 003.jpeg|80px]][[ウミグモ]] |
|label1=[[ウミグモ綱]]|1=[[File:Callipallene brevirostris (YPM IZ 077244) 003.jpeg|80px]][[ウミグモ]] |
||
|label2=<small>[[真鋏角類]]</small>|2={{clade |
|label2=<small>[[真鋏角類]]</small>|2={{clade |
||
|label1=[[節口綱]]<br/><small>(非単系統群)</small>|1=[[ファイル:Tachypleus gigas.JPG|90px]][[カブトガニ類|カブトガニ]]、† [[ウミサソリ]]、†[[カスマ |
|label1=[[節口綱]]<br/><small>(非単系統群)</small>|1=[[ファイル:Tachypleus gigas.JPG|90px]][[カブトガニ類|カブトガニ]]、† [[ウミサソリ]]、†[[カスマタスピス類]]など |
||
|label2=[[クモガタ綱]]|2=[[File:Haeckel Arachnida.jpg|80px]][[クモ]]、[[サソリ]]、[[ダニ]]など |
|label2=[[クモガタ綱]]|2=[[File:Haeckel Arachnida.jpg|80px]][[クモ]]、[[サソリ]]、[[ダニ]]など |
||
}} }} |
}} }} |
||
| 340行目: | 342行目: | ||
}} }} }} }} }} }} }} }} }} }} |
}} }} }} }} }} }} }} }} }} }} |
||
{{-}} |
{{-}} |
||
また、節足動物の胚発生において、眼と前大脳 |
また、現生節足動物の[[胚]][[発生学|発生]]において、眼と前大脳が由来する先頭の体節、いわゆる[[先節]]には[[付属肢]]を持たない(もしくは第1触角をもつ)というのが従来の解釈であった。しかし、多くの節足動物の口の前にある蓋状の構造体、いわゆる[[上唇 (節足動物)|上唇]]と[[ハイポストーマ]]は、著しく融合・退化した先節由来の1対の[[付属肢]]であることがのちに有力視されつつある<ref name=":28">{{Cite journal|last=Haas|first=M. Susan|last2=Brown|first2=Susan J.|last3=Beeman|first3=Richard W.|date=2001-03-08|title=Homeotic evidence for the appendicular origin of the labrum in Tribolium castaneum|url=https://doi.org/10.1007/s004270000129|journal=Development Genes and Evolution|volume=211|issue=2|pages=96–102|language=en|doi=10.1007/s004270000129|issn=0949-944X}}</ref><ref name=":29">{{Cite journal|last=Du|first=Xiaoliang|last2=Yue|first2=Chao|last3=Hua|first3=Baozhen|date=2009-08|title=Embryonic development of the scorpionfly ''Panorpa emarginata'' Cheng with special reference to external morphology (Mecoptera: Panorpidae)|url=https://onlinelibrary.wiley.com/doi/abs/10.1002/jmor.10736|journal=Journal of Morphology|volume=270|issue=8|pages=984–995|language=en|doi=10.1002/jmor.10736|issn=0362-2525}}</ref>。この構造体は、[[有爪動物]]の触角や基盤的な節足動物([[ラディオドンタ類]]など)の[[ラディオドンタ類#前部付属肢|前部付属肢]]との[[相同]]性まで議論をなされ、初期の節足動物、果ては[[汎節足動物]]全般における頭部の起源と進化を示唆する手掛かりの1つとして注目される<ref name=":25" /><ref name="stem" /><ref name="head" />。 |
||
{{Main|上唇_(節足動物)#由来|ラディオドンタ類#前部付属肢の対応関係|汎節足動物#起源と進化}} |
|||
{| class="wikitable" style="margin:1em auto 1em auto" |
{| class="wikitable" style="margin:1em auto 1em auto" |
||
|+ |
|+ |
||
| 465行目: | 467行目: | ||
<gallery mode="packed" heights="200"> |
<gallery mode="packed" heights="200"> |
||
ファイル:20191106 Panarthropoda head segments appendages ja.png|2017年現在で広く認められる、様々な[[汎節足動物]]における前部の体節と付属肢の対応関係<ref name="head" />。先節は「0」 |
ファイル:20191106 Panarthropoda head segments appendages ja.png|2017年現在で広く認められる、様々な[[汎節足動物]]における前部の[[体節]]と[[付属肢]]の対応関係<ref name="head" />。[[先節]]は「0」、頭部に含まれる部分は暗灰色、前大脳性・中大脳性・後大脳性の体節と付属肢はそれぞれP(赤)・D(黄)T(緑)で示される。 |
||
</gallery> |
</gallery> |
||
=== 下位分類 === |
=== 下位分類 === |
||
[[ファイル:Eukaryote species pie tree.png|350px|サムネイル|[[真核生物]]の内部系統と2017年までの |
[[ファイル:Eukaryote species pie tree.png|350px|サムネイル|[[真核生物]]の内部系統と2017年までの種数の円グラフ。その半分以上(右下の[[鋏角類]] Chelicerates から左上の大型[[鱗翅類]] Macrolepidoptera まで)が節足動物に含まれるグループである。]] |
||
節足動物は記載された種数の最も多い[[動物]][[門 (分類学)|門]]である<ref name="Daley2018" /><ref name=":1" /><ref name=":2" />。その数は動物の種のほとんどを占めるというだけでなく、[[真核生物]]の大部分をも占める<ref name=":1" /><ref>{{Cite journal|last=Basset|first=Yves|last2=Cizek|first2=Lukas|last3=Cuenoud|first3=Philippe|last4=Didham|first4=Raphael|last5=Guilhaumon|first5=Francois|last6=Missa|first6=Olivier|last7=Novotny|first7=Vojtech|last8=Ødegaard|first8=Frode|last9=Roslin|first9=Tomas|date=2012-12-14|title=Arthropod Diversity in a Tropical Forest|url=https://www.researchgate.net/publication/233904266_Arthropod_Diversity_in_a_Tropical_Forest|journal=Science|volume=338|pages=1481–1484|doi=10.1126/science.1226727}}</ref>。2011年まででは、100万種以上の[[六脚類]]・11万種以上の[[鋏角類]]・6万種以上の[[甲殻類]]・1万種以上の[[多足類]]という計120万種以上の現生節足動物が記載された<ref name="zhang2011" /><ref name=":2" />。また、[[絶滅]]した[[三葉虫]]も大きなグループであり、1万種以上が記載された<ref>{{Cite web|url=https://australianmuseum.net.au/what-are-trilobites|title=What are trilobites? - Australian Museum|accessdate=2018-10-25|website=australianmuseum.net.au|language=en}}</ref>。 |
節足動物は記載された[[種 (分類学)|種]]数の最も多い[[動物]][[門 (分類学)|門]]である<ref name="Daley2018" /><ref name=":1" /><ref name=":2" />。その数は動物の種のほとんどを占めるというだけでなく、[[真核生物]]の大部分をも占める<ref name=":1" /><ref>{{Cite journal|last=Basset|first=Yves|last2=Cizek|first2=Lukas|last3=Cuenoud|first3=Philippe|last4=Didham|first4=Raphael|last5=Guilhaumon|first5=Francois|last6=Missa|first6=Olivier|last7=Novotny|first7=Vojtech|last8=Ødegaard|first8=Frode|last9=Roslin|first9=Tomas|date=2012-12-14|title=Arthropod Diversity in a Tropical Forest|url=https://www.researchgate.net/publication/233904266_Arthropod_Diversity_in_a_Tropical_Forest|journal=Science|volume=338|pages=1481–1484|doi=10.1126/science.1226727}}</ref>。2011年まででは、100万種以上の[[六脚類]]・11万種以上の[[鋏角類]]・6万種以上の[[甲殻類]]・1万種以上の[[多足類]]という計120万種以上の現生節足動物が記載された<ref name="zhang2011" /><ref name=":2" />。また、[[絶滅]]した[[三葉虫]]も大きなグループであり、1万種以上が記載された<ref>{{Cite web|url=https://australianmuseum.net.au/what-are-trilobites|title=What are trilobites? - Australian Museum|accessdate=2018-10-25|website=australianmuseum.net.au|language=en}}</ref>。 |
||
* '''[[鋏角亜門]]''' '''{{sname||Chelicerata}}''' |
* '''[[鋏角亜門]]''' '''{{sname||Chelicerata}}''' |
||
**[[ウミグモ綱]] (皆脚綱){{sname||Pycnogonida}} - [[ウミグモ]] |
**[[ウミグモ綱]] (皆脚綱){{sname||Pycnogonida}} - [[ウミグモ]] |
||
**[[節口綱]] (腿口綱){{sname|Merostomata}}([[側系統群]]) - [[カブトガニ類]]、†[[ウミサソリ]]、†[[カスマ |
**[[節口綱]] (腿口綱){{sname|Merostomata}}([[側系統群]]) - [[カブトガニ類]]、†[[ウミサソリ]]、†[[カスマタスピス類]]<ref name=":30">{{Cite book|title=ぞわぞわした生きものたち: 古生代の巨大節足動物|url=https://www.worldcat.org/title/zowazowashita-ikimonotachi-koseidai-no-kyodai-sessoku-dobutsu/oclc/816905375|publisher=ソフトバンククリエイティブ|date=2012|location=東京|isbn=978-4-7973-4411-0|oclc=816905375|language=Japanese|first=隆一|last=金子}}</ref>など |
||
**[[クモガタ綱]] (蛛形綱、[[クモ綱]]){{sname||Arachnida}} - [[クモ]]、[[サソリ]]、[[ダニ]]など |
**[[クモガタ綱]] (蛛形綱、[[クモ綱]]){{sname||Arachnida}} - [[クモ]]、[[サソリ]]、[[ダニ]]など |
||
<gallery> |
<gallery> |
||
ファイル:Nymphon-leptocheles.jpg|[[ウミグモ]]の1種 |
ファイル:Nymphon-leptocheles.jpg|[[ウミグモ]]の1種 |
||
ファイル:Origin of Vertebrates Fig 005.png|様々な[[カブトガニ類]] |
ファイル:Origin of Vertebrates Fig 005.png|様々な[[カブトガニ類]] |
||
ファイル:20200607 Chasmataspidida Octoberaspis Hoplitaspis Chasmataspis Diploaspis.png|様々な[[カスマ |
ファイル:20200607 Chasmataspidida Octoberaspis Hoplitaspis Chasmataspis Diploaspis.png|様々な[[カスマタスピス類]] |
||
ファイル:The Eurypterida of New York figure np 1.jpg|様々な[[ウミサソリ]]類 |
ファイル:The Eurypterida of New York figure np 1.jpg|様々な[[ウミサソリ]]類 |
||
ファイル:Jorougumo-1.jpg|[[ジョロウグモ]] |
ファイル:Jorougumo-1.jpg|[[ジョロウグモ]] |
||
</gallery> |
</gallery> |
||
* '''[[多足類|多足亜門]]''' '''{{sname||Myriapoda}}''' |
* '''[[多足類|多足亜門]]''' '''{{sname||Myriapoda}}''' |
||
| 550行目: | 552行目: | ||
;(目)[[:en:Hymenocarina|Hymenocarina]]<ref name=":8" /> |
;(目)[[:en:Hymenocarina|Hymenocarina]]<ref name=":8" /> |
||
:[[カナダスピス]]、[[ブランキオカリス]]、[[オダライア]]、[[ワプティア]]など。真節足動物<ref name="stem" /><ref name="Alacaris" />/[[大顎類]]<ref name=":8" /><ref name=":24" />/[[汎甲殻類]]<ref name=":11">{{Cite journal|last=Vannier|author=|first=Jean|last2=Aria|first2=Cédric|last3=Taylor|first3=Rod S.|last4=Caron|first4=Jean-Bernard|year=2018|title=Waptia fieldensis Walcott, a mandibulate arthropod from the middle Cambrian Burgess Shale|url=https://royalsocietypublishing.org/doi/10.1098/rsos.172206|journal=Royal Society Open Science|volume=5|issue=6|page=|pages=172206|doi=10.1098/rsos.172206|pmid=30110460|pmc=PMC6030330}}</ref>のいずれかのステムグループ。[[カンブリア紀]]に生息。 |
:[[カナダスピス]]、[[ブランキオカリス]]、[[オダライア]]、[[ワプティア]]など。真節足動物<ref name=":34">{{Cite journal|last=Legg|first=David A.|last2=Sutton|first2=Mark D.|last3=Edgecombe|first3=Gregory D.|last4=Caron|first4=Jean-Bernard|date=2012-12-07|title=Cambrian bivalved arthropod reveals origin of arthrodization|url=https://www.researchgate.net/publication/232231696|journal=Proceedings of the Royal Society B: Biological Sciences|volume=279|issue=1748|pages=4699–4704|doi=10.1098/rspb.2012.1958|pmid=23055069|pmc=PMC3497099}}</ref><ref name=":13" /><ref name="stem" /><ref name="Alacaris" />/[[大顎類]]<ref name=":8" /><ref name=":24" />/[[汎甲殻類]]<ref name=":11">{{Cite journal|last=Vannier|author=|first=Jean|last2=Aria|first2=Cédric|last3=Taylor|first3=Rod S.|last4=Caron|first4=Jean-Bernard|year=2018|title=Waptia fieldensis Walcott, a mandibulate arthropod from the middle Cambrian Burgess Shale|url=https://royalsocietypublishing.org/doi/10.1098/rsos.172206|journal=Royal Society Open Science|volume=5|issue=6|page=|pages=172206|doi=10.1098/rsos.172206|pmid=30110460|pmc=PMC6030330}}</ref>のいずれかのステムグループ。[[カンブリア紀]]に生息。 |
||
<gallery> |
<gallery> |
||
ファイル:Canadaspis.jpg|[[カナダスピス]] |
ファイル:Canadaspis.jpg|[[カナダスピス]] |
||
| 559行目: | 561行目: | ||
;[[フーシェンフイア目]] [[:en:Fuxianhuiida|Fuxianhuiida]]<ref name="Alacaris">{{Cite journal|last=Yang|first=Jie|last2=Ortega-Hernández|first2=Javier|last3=Legg|first3=David A.|last4=Lan|first4=Tian|last5=Hou|first5=Jin-bo|last6=Zhang|first6=Xi-guang|date=2018-02-01|title=Early Cambrian fuxianhuiids from China reveal origin of the gnathobasic protopodite in euarthropods|url=https://www.nature.com/articles/s41467-017-02754-z|journal=Nature Communications|volume=9|issue=1|language=En|doi=10.1038/s41467-017-02754-z|issn=2041-1723}}</ref> |
;[[フーシェンフイア目]] [[:en:Fuxianhuiida|Fuxianhuiida]]<ref name="Alacaris">{{Cite journal|last=Yang|first=Jie|last2=Ortega-Hernández|first2=Javier|last3=Legg|first3=David A.|last4=Lan|first4=Tian|last5=Hou|first5=Jin-bo|last6=Zhang|first6=Xi-guang|date=2018-02-01|title=Early Cambrian fuxianhuiids from China reveal origin of the gnathobasic protopodite in euarthropods|url=https://www.nature.com/articles/s41467-017-02754-z|journal=Nature Communications|volume=9|issue=1|language=En|doi=10.1038/s41467-017-02754-z|issn=2041-1723}}</ref> |
||
:[[フーシェンフイア類]]。[[フーシェンフイア]]、[[チェンジャンゴカリス]]など。真節足動物<ref name="stem" /><ref name="Alacaris" />/[[大顎類]]<ref name=":8" /><ref name=":11" /><ref name=":24" />のいずれかのステムグループ。[[カンブリア紀]]に生息。 |
:[[フーシェンフイア類]]。[[フーシェンフイア]]、[[チェンジャンゴカリス]]など。真節足動物<ref name=":34" /><ref name=":13" /><ref name="stem" /><ref name="Alacaris" />/[[大顎類]]<ref name=":8" /><ref name=":11" /><ref name=":24" />のいずれかのステムグループ。[[カンブリア紀]]に生息。 |
||
<gallery> |
<gallery> |
||
ファイル: |
ファイル:20211117 Fuxianhuia protensa.png|[[フーシェンフイア]] |
||
ファイル: |
ファイル:20211117 Chengjiangocaris kunmingensis.png|[[チェンジャンゴカリス]] |
||
ファイル:20211117 Alacaris mirabilis.png|[[アラカリス]] |
|||
ファイル:Guangweicaris restoration.png|[[グァンウェイカリス]] |
|||
</gallery> |
</gallery> |
||
| 576行目: | 580行目: | ||
;[[ブラドリア目]] {{sname||Bradoriida}} |
;[[ブラドリア目]] {{sname||Bradoriida}} |
||
:[[ブラドリア類]]。[[ブラドリア]]、[[クンミンゲラ]]など。真節足動物<ref name=":12">{{Cite journal|last=Zhai|first=Dayou|last2=Williams|first2=Mark|last3=Siveter|first3=David J.|last4=Harvey|first4=Thomas H. P.|last5=Sansom|first5=Robert S.|last6=Gabbott|first6=Sarah E.|last7=Siveter|first7=Derek J.|last8=Ma|first8=Xiaoya|last9=Zhou|first9=Runqing|date=2019-09-03|title=Variation in appendages in early Cambrian bradoriids reveals a wide range of body plans in stem-euarthropods|url=https://www.nature.com/articles/s42003-019-0573-5|journal=Communications Biology|volume=2|issue=1|pages=1–6|language=en|doi=10.1038/s42003-019-0573-5|issn=2399-3642}}</ref>/[[大顎類]]<ref name=":13">{{Cite journal|last=Legg|author=|first=David A.|last2=Sutton|first2=Mark D.|last3=Edgecombe|first3=Gregory D.|year=2013|title=Arthropod fossil data increase congruence of morphological and molecular phylogenies|url=https://www. |
:[[ブラドリア類]]。[[ブラドリア]]、[[クンミンゲラ]]など。真節足動物<ref name=":12">{{Cite journal|last=Zhai|first=Dayou|last2=Williams|first2=Mark|last3=Siveter|first3=David J.|last4=Harvey|first4=Thomas H. P.|last5=Sansom|first5=Robert S.|last6=Gabbott|first6=Sarah E.|last7=Siveter|first7=Derek J.|last8=Ma|first8=Xiaoya|last9=Zhou|first9=Runqing|date=2019-09-03|title=Variation in appendages in early Cambrian bradoriids reveals a wide range of body plans in stem-euarthropods|url=https://www.nature.com/articles/s42003-019-0573-5|journal=Communications Biology|volume=2|issue=1|pages=1–6|language=en|doi=10.1038/s42003-019-0573-5|issn=2399-3642}}</ref>/[[大顎類]]<ref name=":13">{{Cite journal|last=Legg|author=|first=David A.|last2=Sutton|first2=Mark D.|last3=Edgecombe|first3=Gregory D.|year=2013|title=Arthropod fossil data increase congruence of morphological and molecular phylogenies|url=https://www.researchgate.net/publication/257205419|journal=Nature Communications|volume=4|issue=1|page=|language=en|issn=2041-1723}}</ref>/[[汎甲殻類]]<ref>{{Cite journal|last=Hou|first=Xianguang|last2=Williams|first2=Mark|last3=Siveter|first3=David J.|last4=Siveter|first4=Derek J.|last5=Aldridge|first5=Richard J.|last6=Sansom|first6=Robert S.|date=2010-06-22|title=Soft-part anatomy of the Early Cambrian bivalved arthropods Kunyangella and Kunmingella: significance for the phylogenetic relationships of Bradoriida|url=https://royalsocietypublishing.org/doi/full/10.1098/rspb.2009.2194|journal=Proceedings of the Royal Society B: Biological Sciences|volume=277|issue=1689|pages=1835-1841|doi=10.1098/rspb.2009.2194|pmid=20181565|pmc=PMC2871875}}</ref>のいずれかのステムグループ。[[カンブリア紀]]に生息。 |
||
;(目){{sname||Phosphatocopina}} |
;(目){{sname||Phosphatocopina}} |
||
:{{snamei||Hesslandona}}、{{snamei||Vestrogothia}}など。[[大顎類]]のステムグループ<ref name=":13" />もしくは[[甲殻類]]<ref>{{Cite book|title=Fossils and Strata, Morphology, Ontogeny and Phylogeny of the Phosphatocopina (Crustacea) from the Upper Cambrian Orsten of Sweden.|url=https://www.worldcat.org/oclc/741343594|publisher=Wiley|date=2009|isbn=1-282-11829-3|oclc=741343594|others=Waloszek, Dieter., Maas, Andreas., Muller, Klaus.}}</ref>。[[カンブリア紀]]に生息。 |
:{{snamei||Hesslandona}}、{{snamei||Vestrogothia}}など。[[大顎類]]のステムグループ<ref name=":13" />もしくは[[甲殻類]]<ref>{{Cite book|title=Fossils and Strata, Morphology, Ontogeny and Phylogeny of the Phosphatocopina (Crustacea) from the Upper Cambrian Orsten of Sweden.|url=https://www.worldcat.org/oclc/741343594|publisher=Wiley|date=2009|isbn=1-282-11829-3|oclc=741343594|others=Waloszek, Dieter., Maas, Andreas., Muller, Klaus.}}</ref>。[[カンブリア紀]]に生息。 |
||
;[[マ |
;[[マーレロモルフ綱]] {{sname||Marrellomorpha}} |
||
:[[マ |
:[[マーレロモルフ類]]<ref name=":31" /><ref name=":32" />。[[マーレラ]]、[[ヴァコシニア]]、[[フルカ]]、[[ミメタスター]]など。真節足動物<ref name=":8" />/[[大顎類]]<ref name=":13" /><ref name=":12" />のいずれかのステムグループ。[[カンブリア紀]] - [[デボン紀]]に生息。 |
||
<gallery> |
<gallery> |
||
ファイル:Marrella splendens reconstruction by Haug et al.png|[[マーレラ]] |
ファイル:Marrella splendens reconstruction by Haug et al.png|[[マーレラ]] |
||
| 593行目: | 597行目: | ||
:[[ユーシカルシノイド類]]<ref name=":31" />。[[ユーシカルシヌス]]、[[ヘテロクラニア]]、[[アパンクラ]]など。[[大顎類]]<ref name=":8" />/[[多足類]]<ref>{{Cite journal|last=Edgecombe|first=Gregory D.|last2=Strullu-Derrien|first2=Christine|last3=Góral|first3=Tomasz|last4=Hetherington|first4=Alexander J.|last5=Thompson|first5=Christine|last6=Koch|first6=Markus|date=2020-04-01|title=Aquatic stem group myriapods close a gap between molecular divergence dates and the terrestrial fossil record|url=https://www.pnas.org/content/early/2020/03/31/1920733117|journal=Proceedings of the National Academy of Sciences|language=en|doi=10.1073/pnas.1920733117|issn=0027-8424|pmid=32253305}}</ref>のいずれかのステムグループ。[[カンブリア紀]] - [[三畳紀]]に生息。 |
:[[ユーシカルシノイド類]]<ref name=":31" />。[[ユーシカルシヌス]]、[[ヘテロクラニア]]、[[アパンクラ]]など。[[大顎類]]<ref name=":8" />/[[多足類]]<ref>{{Cite journal|last=Edgecombe|first=Gregory D.|last2=Strullu-Derrien|first2=Christine|last3=Góral|first3=Tomasz|last4=Hetherington|first4=Alexander J.|last5=Thompson|first5=Christine|last6=Koch|first6=Markus|date=2020-04-01|title=Aquatic stem group myriapods close a gap between molecular divergence dates and the terrestrial fossil record|url=https://www.pnas.org/content/early/2020/03/31/1920733117|journal=Proceedings of the National Academy of Sciences|language=en|doi=10.1073/pnas.1920733117|issn=0027-8424|pmid=32253305}}</ref>のいずれかのステムグループ。[[カンブリア紀]] - [[三畳紀]]に生息。 |
||
<gallery> |
<gallery> |
||
ファイル:Euthycarcinus kessleri.jpg|{{snamei||Euthycarcinus}} |
ファイル:Euthycarcinus kessleri.jpg|{{snamei||Euthycarcinus}}(旧復元) |
||
</gallery> |
</gallery> |
||
;[[嚢頭類|嚢頭綱]] {{sname||Thylacocephala}} |
;[[嚢頭類|嚢頭綱]] {{sname||Thylacocephala}} |
||
:[[嚢頭類]]([[ティラコセファルス類]]<ref name=":31" />)。{{snamei||Thylacares}}、[[ドロカリス]]、[[キタカミカリス]]など。[[甲殻類]]としての位置は確実 |
:[[嚢頭類]]([[ティラコセファルス類]]<ref name=":31" />)。{{snamei||Thylacares}}、[[ドロカリス]]、[[キタカミカリス]]([[アンキトカゾカリス]]の一種とされる場合もある<ref>{{Cite journal|last=Laville|first=Thomas|last2=Smith|first2=Christopher P. A.|last3=Forel|first3=Marie-Béatrice|last4=Brayard|first4=Arnaud|last5=Charbonnier|first5=Sylvain|date=2021-02-25|title=REVIEW OF EARLY TRIASSIC THYLACOCEPHALA|url=https://riviste.unimi.it/index.php/RIPS/article/view/15188|journal=RIVISTA ITALIANA DI PALEONTOLOGIA E STRATIGRAFIA|volume=127|issue=1|language=en|doi=10.13130/2039-4942/15188|issn=2039-4942}}</ref>)など。[[甲殻類]]としての位置は不確実<ref>{{Cite journal|last=Lange|first=Sven|last2=Schram|first2=Frederick R.|last3=Steeman|first3=Fedor A.|last4=Hof|first4=Cees H. J.|date=2001|title=New Genus and Species from the Cretaceous of Lebanon Links the Thylacocephala To the Crustacea|url=https://onlinelibrary.wiley.com/doi/abs/10.1111/1475-4983.00207|journal=Palaeontology|volume=44|issue=5|pages=905–912|language=en|doi=10.1111/1475-4983.00207|issn=1475-4983}}</ref>。[[オルドビス紀]]<ref>{{Cite journal|last=Roy|first=Peter Van|last2=Rak|first2=Štěpán|last3=Budil|first3=Petr|last4=Fatka|first4=Oldřich|date=2021|title=Upper Ordovician Thylacocephala (Euarthropoda, Eucrustacea) from Bohemia indicate early ecological differentiation|url=https://onlinelibrary.wiley.com/doi/abs/10.1002/spp2.1363|journal=Papers in Palaeontology|volume=7|issue=3|pages=1727–1751|language=en|doi=10.1002/spp2.1363|issn=2056-2802}}</ref>([[カンブリア紀]]?)- [[白亜紀]]に生息。 |
||
<gallery> |
<gallery> |
||
ファイル:Thylacares brandonensis S12862-014-0159-2-10.jpg| |
ファイル:Thylacares brandonensis S12862-014-0159-2-10.jpg|''[[:en:Thylacares|Thylacares]]'' |
||
ファイル:Dollocaris ingens reconstruction.jpg|[[ドロカリス]] |
ファイル:Dollocaris ingens reconstruction.jpg|[[ドロカリス]] |
||
ファイル:Ostenocaris cypriformis.jpg| |
ファイル:Ostenocaris cypriformis.jpg|''[[:en:Ostenocaris|Ostenocaris]]'' |
||
ファイル:Proetus concinnus reconstruction.JPG| |
ファイル:Proetus concinnus reconstruction.JPG|''[[:en:Ainiktozoon|Ainiktozoon]]''(上、青) |
||
</gallery> |
</gallery> |
||
;(目){{sname||Cyclida}} |
;(目){{sname||Cyclida}} |
||
{{snamei||Cyclus}}、{{snamei||Schramine}}、{{snamei||Americlus}}など。[[甲殻類]]としての位置は確実 |
{{snamei||Cyclus}}、{{snamei||Schramine}}、{{snamei||Americlus}}など。[[甲殻類]]としての位置は不確実<ref>{{Cite journal|last=Schädel|first=Mario|last2=Haug|first2=Joachim T.|date=2020/05|title=A new interpretation of the enigmatic fossil arthropod Anhelkocephalon handlirschi Bill, 1914 – important insights in the morphology of Cyclida|url=https://bioone.org/journals/Palaeodiversity/volume-13/issue-1/pale.v13.a7/A-new-interpretation-of-the-enigmatic-fossil-arthropod-Anhelkocephalon-handlirschi/10.18476/pale.v13.a7.full|journal=Palaeodiversity|volume=13|issue=1|pages=69–81|doi=10.18476/pale.v13.a7|issn=1867-6294}}</ref>。[[石炭紀]] - [[白亜紀]]に生息。 |
||
<gallery> |
<gallery> |
||
File:Cyclus americanus 34.JPG|{{snamei||Cyclus americanus}}(化石) |
File:Cyclus americanus 34.JPG|{{snamei||Cyclus americanus}}(化石) |
||
2021年12月26日 (日) 06:58時点における版
| 節足動物 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 現生および絶滅した様々な節足動物[注釈 1]
| ||||||||||||||||||
| 地質時代 | ||||||||||||||||||
| カンブリア紀 - 現世 | ||||||||||||||||||
| 分類 | ||||||||||||||||||
| ||||||||||||||||||
| 学名 | ||||||||||||||||||
| Arthropoda von Siebold, 1848[2][3] | ||||||||||||||||||
| 和名 | ||||||||||||||||||
| 節足動物 | ||||||||||||||||||
| 英名 | ||||||||||||||||||
| Arthropod | ||||||||||||||||||
| 亜門 | ||||||||||||||||||
|
絶滅群は本文参照 |
節足動物(せっそくどうぶつ、英: Arthropod、学名:Arthropoda[2])とは、昆虫類・甲殻類・クモ類・ムカデ類など、外骨格と関節を持つ動物を含んだ分類群。分類学上は節足動物門とされる。動物界最大かつ多様性の最も高い動物門であり[4][1][5][6]、現生種は全動物種の85%以上を占め、約110万種が記載される[7]。陸・海・空・土中・寄生などあらゆる場所に進出し、様々な生態系と深く関わっている。なお、いわゆる「虫」の範疇に入る動物は当動物門のものが多い[注釈 2]。
学名「Arthropoda」はギリシア語の「άρθρον」(arthron, 関節)と「πούς」(pous, 脚)の合成語であり、本動物門の関節に分かれた付属肢(関節肢)に因んで名づけられた[8]。
形態学、解剖学と生理学
体節制

節足動物の形態は多様で、分類群によって様々な外見を持つ。体の表面はキチン質とタンパク質等からなるクチクラ(cuticle)でできた外骨格(exoskeleton)で覆われる。成長に伴い体のサイズが大きくなるときには、脱皮により古い外骨格は脱ぎ捨てられ、新しい外骨格が形成される。節足動物は体節制(segmentation)をもち、すなわち体は体節(somite)という単位の繰り返し構造からなり、各体節は原則として1対の付属肢をもつ(後述)[9]。体の背腹は、それぞれ背板(tergite, tergum)と腹板(sternite, sternum)という外骨格に覆われており、体節の両側で更に側板(pleuron)を持つ場合もある。表面を被うこれらの外骨格も体節単位になっており、体節の間は関節状に可動であることが多い。体の先頭の体節は先節(ocular somite、古典的には口前葉 acron)といい、節足動物の眼と口はここに由来する[10]。一部の分類群は、体の末端に尾節(telson)という尾のような構造もある[9]。



ただし、節足動物は異規体節制(Heteronomous metamerism[11])がある程度以上発達し、複数の体節が組み合わされ、外観上あるいは機能上の単位を構成する(tagmosis、tagmatization)[9]例が多く見られる。例えば、体を「頭部・胸部・腹部」「頭部・胸部・尾部」「前体・中体・終体」などの3部、または「頭部・胴部」「頭胸部・腹部」「前体・後体」などの2部に分けて呼ぶ場合があり、これは節足動物の各分類群ごとの特徴として用いられる[9]。特に前方の合体節(頭部・頭胸部・前体)は往々にして体節の癒合が進み、外見上では元の体節構造が見当たらず、すべて単一の外骨格に覆われている[9]。
なお、一部の寄生性の分類群には極端に特化し、外見上の体節構造が全く見当たらない例もある[9]。
付属肢


各体節からは、原則として1対の関節肢(arthropodized appendage)という本群に特有の付属肢が出ている。これが「節足動物」という名前およびその学名の由来となっている[8]。関節肢も体と同様に外骨格で覆われ、関節によって分かれた肢節(podomere)からなる。関節肢は所属する分類群や位置によって歩脚・遊泳脚・鋏・鎌・顎・触角・鰓・生殖肢など様々な機能に応じて色んな形に特化している[9]。例えば頭部には感覚用の触角と摂食用の顎、胴部には移動用の歩脚を持つなど、節足動物は、往々にして異なる機能を担った様々な関節肢を掛け合わせて、「アーミーナイフのように、独自の機能を有する複数の道具が同時にセットされる」とも比喩される[12]。また、節足動物は多くが口の直前に上唇(labrum)もしくはハイポストーマ(hypostome)と呼ばれる1枚の蓋状の構造体があり、これも付属肢由来の部分ではないかと思われる[13][14][15][10]。なお、前述の体節のように、寄生性甲殻類や昆虫の幼虫の中には、関節肢が不明瞭もしくは完全に退化消失した例もある[9]。
-
 軟甲類の体制模式図:頭部には2対の触角と3対の顎・胸部には8対の胸肢・腹部には5対の腹肢と1対の尾肢をもつ
軟甲類の体制模式図:頭部には2対の触角と3対の顎・胸部には8対の胸肢・腹部には5対の腹肢と1対の尾肢をもつ -
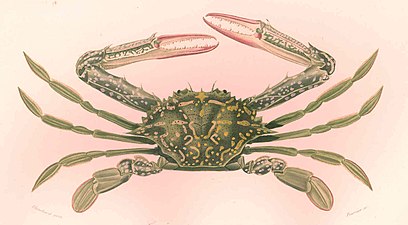 鋏型の第1脚、歩脚型の第2-4脚とヘラ状の第5脚(遊泳脚)をもつタイワンガザミ(ワタリガニ)
鋏型の第1脚、歩脚型の第2-4脚とヘラ状の第5脚(遊泳脚)をもつタイワンガザミ(ワタリガニ)



関節肢は、内肢(endopod)と外肢(exopod)もしくは外葉(exite、副肢 epipod)をもつ二叉型付属肢(二肢型付属肢、biramous appendage)が基本だと思われる[16][17]。しかし、現生節足動物の中でこの性質を明瞭に受け継いだのは多くが甲殻類で[18]、他の分類群においては内肢しか見られない単枝型付属肢(単肢型付属肢、uniramous appendage)がほとんどである。また、三葉虫とメガケイラ類をはじめとする多くの古生代の絶滅分類群は、胴部の付属肢がヒレ状の外肢と歩脚状の内肢でできた二叉型である[16][17]。
![三葉虫の二叉型の歩脚]]](/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Trilobite_ventral_appendages_IT.PNG)





運動
関節の隙間は柔らかい節間膜(arthrodial membrane)に覆われており、2つの関節の外骨格内側に付着する筋肉を介して運動をする。胴部の関節は様々な方向へ湾曲できることが多いが、関節肢の関節は往々にして1対の蝶番のような構造、いわゆる関節丘(ピボット、pivot)に固定されて一つの平面上でしか動かない。そのため節足動物の関節肢、特に基部は往々にして複数の節に分かれて、様々な動きに対応できるようになる[19]。また、外骨格は体の内側へ延長し、いわゆる腱(内突起、internal tendon, apodeme)となり、筋肉に付着面を提供することが普通である[20]。例えばカニの鋏の中では、可動指の関節に繋がった、大量の筋肉が付着する板状の腱が見られる。また、筋肉の代わりに体中の血リンパ(後述)の水圧を調整することで関節肢を動かす例もある[21]。
神経系と循環系


P: 前大脳、D: 中大脳、T: 後大脳、CV: 腹髄神経索
多くの左右相称動物と同様、節足動物は体腔を持ち、消化管は体の前後に貫通し、いわゆる口と肛門という2つ開口を持つ。体節に貫通する紐状の心臓と中枢神経はそれぞれ体の背面と腹面に付く。
循環系は開放血管系(open circulatory system)であり、細胞外液はリンパ液や血液という区別は存在せず、リンパ液と血液の役割を兼ねている血リンパ(hemolymph)が心臓と組織の間隙(血体腔)に流れる。心臓の伸縮や体の運動によって血リンパは心臓の動脈から体の静脈と呼吸器などの器官を通り、心門を通じて再び心臓に戻る。血リンパの中には免疫系に関わる血球がある。
体節制をもつ他の前口動物に似て、節足動物の神経系の様式ははしご形神経系(ladder-like nervous system)である。腹側(ただし脳は背側[1])の1対の神経索が体の前後を走り、各体節に由来する神経節(ganglion)は、左右の連絡(横連合、commissure)で繋がっている。
体の前端部には脳があり、食道の前上方にあることから食道上神経節(supraoesophageal ganglion、大脳神経節[22])とも呼ぶ。この脳は先頭3つの体節(先節・第1体節・第2体節)と共に3対の神経節が融合した結果であり、前大脳(protocerebrum)・中大脳(deutocerebrum)・後大脳(tritocerebrum)という3つの脳神経節から構成される[23][15][10]。前大脳には複眼からの視覚情報を処理する視葉(optic lobe)、嗅覚の識別や記憶および感覚神経の統御を司るキノコ体(mushroom body)、視覚行動の統御を行う中心複合体(central body)を持つ[23]。脳は前大脳をはじめとして背側にあるため、中央もしくは直後から食道を囲み、食道神経環(circumesophageal nerve ring)を介して腹側の腹神経索(ventral nerve cord)に連結する。昆虫・甲殻類などの大顎類の場合、食道神経環の直後は顎(大顎・小顎/下唇)に対応する神経節で、まとめて食道下神経節(suboesophageal ganglion、顎神経節[22])といい[23]、ハエやハチ、チョウなどにおいては脳と融合し頭部神経節を構成する(この場合は食道上神経節のみならず、食道下神経節も脳の一部と扱う)[23]。一部の体節の融合が進み、神経が集中してはしご形が不明瞭な場合もあり、例えばカブトガニやクモガタ類の前体において脳と腹神経索を集約させた synganglion、およびカニや派生的な昆虫において著しく集約した胸部と腹部の神経節がその例である[24]。
感覚器

節足動物は様々な感覚器を通じて周りの環境を感知する。体表は常に剛毛(刺毛、感覚毛、setae)をもち、種によっては触覚・振動・音・水流や気流・温度・匂いや味・化学物質など視力以外の感覚を持つ。鋏角類以外の節足動物の頭部は、往々にして触角(Antenna)という関節肢をもち、ほとんどの場合は重要な感覚器官である。なお、触角のない鋏角類の中でも、ウデムシやサソリモドキの様に一部の歩脚が感覚器官に特化した例がある[25]。

他にも昆虫の小顎と下唇にある顎鬚(palp)が嗅覚や味覚に関わり、サソリの櫛状板・ヒヨケムシのラケット器官・一部の昆虫と甲殻類の後端にある尾毛(cercus、caudal ramus)が感覚器官として用いられるほか、コオロギやキリギリスが前脚に[26]・バッタが腹部に[27]・カマキリが後胸部腹面[28]に有する、特殊化した聴覚器官である鼓膜器官が知られる。
-
 長い触角と尾毛をもつシミ
長い触角と尾毛をもつシミ -
 Zabalius aridus(キリギリス科)の前脚に位置する鼓膜
Zabalius aridus(キリギリス科)の前脚に位置する鼓膜 -
 触角のような第1脚をもつウデムシ
触角のような第1脚をもつウデムシ -
 サソリの櫛状板(黄色、赤印先)
サソリの櫛状板(黄色、赤印先)




眼

)_(36522833083).jpg)
節足動物は、中眼(median eye)と側眼(lateral eye)という先節に由来する[10]2種類の眼を持ち、その中で中眼は単眼(ocellus, simple eye)で、側眼は複眼(compound eye)であることが基本と思われる[29][30]。しかしその片方しか持たず、複眼が単眼(側単眼)と化したり、更に祖先形質として眼をもつ多くの動物の分類群と同様、眼が二次的に退化消失した節足動物もある。
複眼は図形認識能力をもち、数多くの個眼(ommatidium)というレンズ様の構成単位からモザイク画のような視覚を形成する。単眼は主に明暗を感知するなど補助的な機能を担うが、一部のクモの単眼は優れた図形認識能力を持ち、例えばハエトリグモの視力は発達しており、内部の網膜をも動かすことができる[21]。
眼をもつ節足動物の中で、六脚類と甲殻類は基本的に中眼と側眼をあわせもつ。後者の場合、中眼はノープリウス幼生期のノープリウス眼に当たる[29]。多足類は全て中眼を欠き、中でゲジ類のムカデは側眼が複眼のままで、他のものは複眼が側単眼に変化した。鋏角類の中でウミグモは中眼のみ、カブトガニとウミサソリは複眼と中眼の両方、現生のクモガタ類は複眼を持たず、中眼と複眼由来の側単眼をあわせもち、或いは片方のみを持つ[30]。また、カンブリア紀の化石節足動物をはじめとし、三葉虫類や基盤的な節足動物と思われるラディオドンタ類も発達した複眼を持つ[31][32][33][34]。






呼吸
節足動物は様々な生息環境に進出しており、それに応じた多様な呼吸様式がみられる。陸生種では気管(trachae)や書肺(book lung)、水生種では鰓(gill)をもつものがある。呼吸器官を持たず、体表でガス交換を行う種類もある。
- 六脚類は主に陸生で発達した気管を持ち、胸部と腹部のそれぞれの節に一対の気門を持つ。水生昆虫の中で一部の幼虫は、水中呼吸に用いられる鰓を持つ。
- 多足類は全て陸上性で、六脚類と似たような気管と気門で呼吸する。
- 鋏角類の中で、水生のカブトガニ類は後体の蓋板にある書鰓で呼吸をし、陸生のクモガタ類は主に書肺や気管(気門)を通じて行う。なお、ウミグモ・コヨリムシと一部のダニは呼吸器官を持たず、体表で直接的にガス交換を行う。
- 甲殻類は通常、付属肢における外葉などの附属体が鰓となって水中に呼吸する[18]。陸生のワラジムシ亜目は腹肢にある白体(偽気管)で呼吸し、ヤシガニの鰓室は陸上での呼吸に用いられる無数の突起物がある[35]。




.jpeg)
繁殖と発育
求愛行動・交尾或いは交接・メイトガード・護卵など、節足動物は分類群によって様々な繁殖行動を持つ。原則として有性生殖を行う卵生動物であるが、サソリやアブラムシなどから単為生殖と卵胎生の例も知られる。

成長に伴い古い外骨格の下で新しい外骨格を形成し、脱皮(Ecdysis)を通じて古い外骨格から抜け出して成長する。新しい外骨格は柔らかく、固くなるのも時間が掛かり、脱皮直後の節足動物は無防備である。そのため節足動物の脱皮は常に隠れ場所で行うことが多い。中でも古い外骨格を摂食して栄養を回収する種類や、一部のクモは脱皮直後のメスを狙って交接することが知られる[21]。
節足動物の幼生は基本的に成体と似たような外見を持つが、甲殻類のノープリウス幼生や昆虫の幼虫など、成長の過程で著しく形態が変化する変態(metamorphosis)を行う分類群も少なくない。昆虫の成虫になる脱皮過程は羽化(Eclosion)と呼ばれる。一部の節足動物、例えば多足類の中には、成体になるまで脱皮に伴って体節と脚の数を増やせ、いわゆる増節変態(anamorphic development)を行うものがある[36]。
-
ハエトリグモの求愛ダンス
-
 護卵するムカデ
護卵するムカデ -
 幼生を背負するサソリ
幼生を背負するサソリ -

-
 羽化直後のアブラゼミ成虫
羽化直後のアブラゼミ成虫 -

-
 ヒョウモンドクチョウの生活環
ヒョウモンドクチョウの生活環






他の動物門との関係性
| |||||||||||||||||||||||||||||||||
| 節足動物の系統的位置 |






節足動物と他の動物門の類縁関係は長らく議論をなされ、20世紀末と2000年代を介して劇的に更新された[37][38][15][39][4]。21世紀以前では、前口動物で体節制を持つなどの共通点から、節足動物と環形動物は近縁である同時に、舌形動物(シタムシ)・有爪動物(カギムシ)・緩歩動物(クマムシ)という3つの動物門は、両者の中間形態を示唆するという考えが主流であった。これらの動物群は、体節動物(Articulata[40])という単系統群を構成すると考えられ[41][42][43][44]、中でも有爪動物と緩歩動物は節足動物に内包され[2][45]、もしくは舌形動物・有爪動物・緩歩動物がまとめて側節足動物(Parathropoda[46])として区別される経緯すらあった[47]。
しかし21世紀以降では、分子系統学をはじめとする多方面(遺伝子発現・解剖学・発生学)の進展により、環形動物は他の体節動物に類縁でなく、むしろトロコフォア幼生を共有する軟体動物などと共に単系統群の冠輪動物(Lophotrochozoa)に属するものであると判明した。同時に他の体節動物も環形動物より、むしろ脱皮などの性質を共有する線形動物などと単系統群になると判明し、脱皮動物(Ecdysozoa[48])として区別されるようになった[49][37][38][39][4]。かつて体節動物の根拠とされてきた環形動物と他の体節動物の体節制も、後に発生学と遺伝子発現の違いにより別起源(収斂進化)であることが示唆される[50][51][52]。更に、かつて側節足動物ともされてきた群の中で、緩歩動物と有爪動物は節足動物に内包されない独立の動物門として広く認められる一方、舌形動物は独立した動物門ではなく、鰓尾類に近縁の甲殻類、すなわち極端に特化した節足動物の一員だと判明した[53][54][4]。
その結果、古典的な「体節動物」と「側節足動物」はいずれも系統関係を反映できない多系統群として解体され、徐々に21世紀以降の分類体系から廃止された[39][4]。21世紀、特に2000年代後期以降では、脱皮動物の中で、節足動物・有爪動物・緩歩動物という3動物門が単系統群を構成する説が広く認められ、まとめて汎節足動物(Panarthropoda[55])と呼ばれている[56][57][58][4]。
なお、脱皮動物と汎節足動物の単系統性が広く認められるものの、汎節足動物と他の脱皮動物(環神経動物 Cycloneuralia[59][60])の系統関係ははっきりしておらず、汎節足動物内の3動物門は形態の類似と分子系統解析の食い違いにより、お互いの系統関係は諸説に分かれている[61]。これらの議論の詳細については汎節足動物#系統関係および汎節足動物#内部系統関係を参照のこと。
.jpg)




起源
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 節足動物の初期系統を中心とする汎節足動物の系統関係[62] (†:絶滅群、青枠:基盤的な節足動物、*:葉足動物) |







知られる最古の節足動物は、およそ5億年前のカンブリア紀に遡る[1]。この地質時代からは、アノマロカリスをはじめとするラディオドンタ類(アノマロカリス類)やオパビニアなど、節足動物的な形質と節足動物らしからぬ形質を掛け合わせた古生物が見つかり、節足動物の初期系統を表したものとして広く注目される(後述)[63][64][33][65][4][66]。しかし、派生的な節足動物の最古の化石記録はそれ以上に早期(体の化石は約5億2,100万年前まで、生痕化石は約5億3,700万年前まで)のため、節足動物の初期系統は中間化石記録を欠いており、実際の起源は記録以上に古かったことが示唆される[1]。なお、カンブリア紀とその直前のエディアカラ紀の化石産地の比較により、その起源は、エディアカラ紀晩期(約5億5,000万年前)より古くはなかったと考えられる[1]。



節足動物の初期系統(ステムグループ)に含まれる古生物、いわゆる基盤的な節足動物(ステム節足動物)は、主にラディオドンタ類、オパビニア、シベリオン類、およびこれらの古生物の特徴を掛け合わせたようなケリグマケラとパンブデルリオンがある。これらの古生物は先頭に特化した1対の前部付属肢をもつことが特徴的で[15][10]、体の両筋に鰭をもつ群(シベリオン類以外の群)は「dinocaridids」(Dinocaridida)、パンブデルリオンとケリグマケラは便宜的に「gilled lobopodians」としてまとめられる場合もある[67][68]。
これらの古生物は少なくとも胴部が柔軟で、一見では節足動物らしくないが、ある程度の節足動物的性質をもつことが分かる[15][10]。いずれも早期の節足動物において特徴的な消化腺をもち[69]、中でもラディオドンタ類は最も節足動物的で、関節肢・複眼・背面の外骨格などという節足動物として決定的な特徴が頭部に出揃っている[15][10][29][4]。オパビニアは節足動物に似た後ろ向きの口と分化した眼を有し[15][10][62]、ラディオドンタ類、パンブデルリオンとケリグマケラに見られる背腹2種類の付属肢要素(背側の鰓/鰭と腹側の鰭/脚)も、節足動物の二叉型付属肢を彷彿とさせる[63][65]。
一方、シベリオン類は姿そのものがれっきとした葉足動物(脚のある蠕虫様の化石汎節足動物)であり、パンブデルリオンとケリグマケラも葉足動物に似た表皮構造と付属肢(葉足)をもつ[63]。このような中間型生物の存在により、汎節足動物の中で、節足動物は有爪動物や緩歩動物と同様、葉足動物から派生した動物群であることが示唆される[15][10][1]。また、ラディオドンタ類の性質により、節足動物の複眼と関節肢の進化(arthropodization)は、胴部の外骨格と関節の進化(arthrodization)より先であったことも示される[15][10][4]。



基盤的な節足動物より派生的で、現生群をも含んだ残り全ての節足動物は一般に真節足動物(Euarthropoda, Deuteropoda)として区別される[15][10]。この系統群をはじめとする節足動物は、頭部は複数体節の融合でできて、胴部まで外骨格と関節肢を備わっており、前述の基盤的な節足動物とは明らかに異なる[15][10]が、キリンシアという、基盤的な節足動物に似た特徴が顕著に見られる中間型生物もある[62]。他にはイソキシス類(イソキシスなど)・メガケイラ類(大付属肢節足動物)・フーシェンフイア類・Hymenocarina類(ワプティア、カナダスピスなど)が挙げられており、それぞれ文献によって現生節足動物の系統群(鋏角類・多足類・甲殻類・六脚類)全体よりも基盤的で[10][15]、もしくは特定の現生群の初期系統を表したものと考えられる(例えばメガケイラ類は基盤的な鋏角類、Hymenocarina類は多足類・甲殻類・六脚類を含んだ系統群に近いともされる)[4][66][62]。
分類
系統関係と体節の相同性
現生の節足動物は、鋏角亜門(クモ、サソリ、カブトガニなど)・多足亜門(ムカデ、ヤスデなど)・甲殻亜門(カニとエビ、フジツボ、ミジンコなど)・六脚亜門(昆虫、トビムシなど)という4つの亜門に分類されている。化石種まで範囲を広げれば、三葉虫などを含んだArtiopoda亜門という過去の大グループや、確定的に現生の亜門に収まらない分類群も数多く知られている(後述参照)[4][66]。
それそれの分類群で特に注目される特徴は、先頭複数体節の融合でできた頭部(真鋏角類の場合は前体)の付属肢である。その基本構成は各亜門によって異なり、次の通りである:
- 六脚類:触角1対・大顎1対・小顎1対・下唇1枚(1対の小顎から癒合した部分)
- 多足類:触角1対・大顎1対・小顎2対
- 甲殻類:触角2対・大顎1対・小顎2対
- 鋏角類(真鋏角類):鋏角1対・触肢1対・脚4対
- 鋏角類(ウミグモ類):鋏肢1対・触肢1対・担卵肢1対・脚1対
- Artiopoda類:触角1対・脚3対以上
これらの付属肢の対応関係(相同性)は、節足動物の内部系統や他の汎節足動物との関係性を大きく左右する指標とされ、そこから繰り広げた議論は「Arthropod head problem」として知られている[10][4]。


![2017年現在で広く認められる、様々な節足動物における前部の体節と付属肢の対応関係[10][25]。頭部および前体に含まれる体節は暗灰色で、前大脳性・中大脳性・後大脳性の体節と付属肢はそれぞれP(赤)・D(黄)T(緑)で示される。](/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:20190913_Arthropoda_head_segments_appendages_ja.png)
21世紀以前の古典的な分類学では、鋏角類の触角を欠くという特徴は二次的退化と考えられ[70]、多足類と六脚類は近縁とされてきた[71]。しかしこれらの知見は、後に発生学・神経解剖学・遺伝子発現・分子系統学など多方面の情報に否定され、代わりに鋏角類の鋏角は大顎類の第1触角に相同で[72][73][74][75]、多足類と六脚類の直接の関連も支持されず、むしろ六脚類は側系統群の甲殻類から分岐したことが強く示唆される(汎甲殻類仮説)[76][77][78][79][80][81][82]。また、通常では鋏角類とされるウミグモ類の系統位置がしばしば疑問視されるようになり、分子系統解析では鋏角類であることを支持するものが多い[83][84][66]が、2000年代ではそれ以外の節足動物と対置すべき説もある[85][79][86]。
三葉虫類をはじめとして、幾つかの絶滅した節足動物の分類群と現生分類群の類縁関係については、未だに定説がない[4]。例えば三葉虫などを含むArtiopoda類は、鋏角類に類縁という伝統的な系統仮説はあった[70]が、頭部の構成に基づいて触角をもつ現生群(多足類・甲殻類・六脚類)に近い[87][88][89]、またはどの現生群よりも基盤的[90][4]とも考えられる。メガケイラ類に関しては、付属肢と脳の構造に基づいて鋏角類に近い[91]、もしくはどの現生群よりも基盤的との説がある[15][10][4]。
こうした研究の発展に伴い、節足動物の高次系統に対して様々な系統仮説が与えられており、以下の例が挙げられる[92][93]。
- 大顎類 Mandibulata
- 多足類・甲殻類・六脚類からなる。大顎を持つことが共有形質とされる。
- 汎甲殻類 Pancrustacea(=八分錘類 Tetraconata)
- 甲殻類と六脚類からなる。複眼の八分割される硝子体などが共有形質とされる。
- 多足鋏角類 Myriochelata(=矛盾足類 Paradoxopoda)
- 多足類と鋏角類からなる。
- 裂肢類 Schizoramia(='CCT' clade)
- 甲殻類・鋏角類・Artiopoda類などからなる。ニ叉型付属肢が共有形質とされる[94]。
- 広義の単肢類 Uniramia sensu lato
- 有爪動物・多足類・六脚類からなる。単枝型付属肢が共有形質とされる。またこの系統仮説に従えば、有爪動物は節足動物に含まれ、もしくは節足動物が多系統群になる。
- Arachnomorpha(=蛛形様類 Arachnata[95])
- Artiopoda類と鋏角類からなる。
- Antennulata
- Artiopoda類・多足類・甲殻類・六脚類からなる。中大脳性(第1体節由来)の第1触角を持つことが共有形質とされる。
- 板肢類 Lamellipedia[94][95]
- Artiopoda類とMarrellomorpha類からなる。
2010年代以降では、少なくとも節足動物は有爪動物を含まない単系統群であることや、大顎類説、汎甲殻類説、およびウミグモ類が鋏角類に含まれる説が広く認められる[4][66]。
| 節足動物の内部系統関係 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 節足動物の各亜門(太字)の類縁関係とその内部高次系統をまとめた分岐図。青い枠以内の分類群、すなわち六脚亜門以外の汎甲殻類は側系統の甲殻亜門に属する。系統位置が不確実なものは、二分岐ではなく多分岐として描いている。亜門が未定/不確実で、もしくはそのクラウングループに含まれていない化石群はここに示さない。 |
_003.jpeg)





.jpg)


.jpeg)

_001.jpeg)


また、現生節足動物の胚発生において、眼と前大脳が由来する先頭の体節、いわゆる先節には付属肢を持たない(もしくは第1触角をもつ)というのが従来の解釈であった。しかし、多くの節足動物の口の前にある蓋状の構造体、いわゆる上唇とハイポストーマは、著しく融合・退化した先節由来の1対の付属肢であることがのちに有力視されつつある[13][14]。この構造体は、有爪動物の触角や基盤的な節足動物(ラディオドンタ類など)の前部付属肢との相同性まで議論をなされ、初期の節足動物、果ては汎節足動物全般における頭部の起源と進化を示唆する手掛かりの1つとして注目される[33][15][10]。
分類群
|
先節 (前大脳) |
1 (中大脳) |
2 (後大脳) |
3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|---|
| 六脚類 | 上唇 * | 触角 * | (退化)* | 大顎 * | 小顎 * | 下唇 * | 脚 |
| 甲殻類 | 上唇 * | 第1触角 * | 第2触角 * | 大顎 * | 第1小顎 * | 第2小顎 * | 脚 |
| 多足類 | 上唇 * | 触角 * | (退化)* | 大顎 * | 第1小顎 * | 第2小顎 * | 脚 |
| 鋏角類(真鋏角類) | 上唇 * | 鋏角 * | 触肢/脚 * | 脚 * | 脚 * | 脚 * | 脚 * |
| 鋏角類(ウミグモ類) | ? * | 鋏肢 * | 触肢 * | 担卵肢 * | 脚 * | 脚 | 脚 |
| Artiopoda類 | ハイポストーマ * | 触角 * | 脚 * | 脚 * | 脚 * | 脚 | 脚 |
| メガケイラ類 | ハイポストーマ * | 大付属肢 * | 脚 * | 脚 * | 脚 * | 脚 | 脚 |
| フーシェンフイア類 | ハイポストーマ * | 触角 * | SPAs * | 脚 | 脚 | 脚 | 脚 |
| ラディオドンタ類[注釈 3] | 前部付属肢 * | 鰭 | 鰭 | 鰭 | 鰭 | 鰭 | 鰭 |
| 緩歩動物[注釈 3] | 歯針 * | 葉足 | 葉足 | 葉足 | 葉足 | - | - |
| 有爪動物[注釈 4] | 触角 * | 顎 * | 粘液腺 | 葉足 | 葉足 | 葉足 | 葉足 |
| 葉足動物[注釈 3] | 触角/前部付属肢 * | 葉足 | 葉足 | 葉足 | 葉足 | 葉足 | 葉足 |
![2017年現在で広く認められる、様々な汎節足動物における前部の体節と付属肢の対応関係[10]。先節は「0」、頭部に含まれる部分は暗灰色、前大脳性・中大脳性・後大脳性の体節と付属肢はそれぞれP(赤)・D(黄)T(緑)で示される。](/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:20191106_Panarthropoda_head_segments_appendages_ja.png)
下位分類

節足動物は記載された種数の最も多い動物門である[1][5][6]。その数は動物の種のほとんどを占めるというだけでなく、真核生物の大部分をも占める[5][97]。2011年まででは、100万種以上の六脚類・11万種以上の鋏角類・6万種以上の甲殻類・1万種以上の多足類という計120万種以上の現生節足動物が記載された[3][6]。また、絶滅した三葉虫も大きなグループであり、1万種以上が記載された[98]。







_crop.jpg)
- 甲殻亜門 Crustacea(側系統群)
- 貝形虫綱 Ostracoda - 貝虫(カイミジンコ)
- ヒゲエビ亜綱 Mystacocarida - ヒゲエビ
- ウオヤドリエビ綱[96](イクチオストラカ綱[100]) Ichthyostraca
- 鰓尾亜綱 Branchiura - 鰓尾類(チョウ、ウオジラミ)
- 舌形亜綱 Pentastomida - シタムシ
- 軟甲綱 Malacostraca - シャコ、カニ、エビ、オキアミ、ダンゴムシなど
- 六幼生綱[96](六齢ノープリウス綱[100]) Hexanauplia
- 鰓脚綱 Branchiopoda - アルテミア、ホウネンエビ、カブトエビ、カイエビ、ミジンコなど
- カシラエビ綱 Cephalocarida - カシラエビ
- ムカデエビ綱 Remipedia - ムカデエビ







絶滅した分類群
以下は上記の現存群(鋏角類・多足類・甲殻類・六脚類)のクラウングループに含まれていない(すなわち該当現存群に対して基盤的な化石分類群、ステムグループ)、もしくは所属する現存群が不確実である化石分類群を取り上げる。
- ラディオドンタ目(放射歯目)Radiodonta - ラディオドンタ類[101][102][103][104](放射歯類[99]、広義のアノマロカリス類[105][106])。アノマロカリス、アンプレクトベルア、ペイトイア、フルディアなど。
-

-

-

-
 様々なラディオドンタ類
様々なラディオドンタ類




- (目)Hymenocarina[90]
- カナダスピス、ブランキオカリス、オダライア、ワプティアなど。真節足動物[107][108][15][16]/大顎類[90][62]/汎甲殻類[109]のいずれかのステムグループ。カンブリア紀に生息。



- フーシェンフイア目 Fuxianhuiida[16]
- フーシェンフイア類。フーシェンフイア、チェンジャンゴカリスなど。真節足動物[107][108][15][16]/大顎類[90][109][62]のいずれかのステムグループ。カンブリア紀に生息。


- メガケイラ綱 Megacheira[94]
- メガケイラ類[101](大付属肢節足動物、大付属肢型節足動物)。ヨホイア、ハイコウカリス、レアンコイリア、フォルティフォルケプスなど。真節足動物[15][16]/鋏角類[91][62]のいずれかのステムグループ。カンブリア紀 - デボン紀に生息。





- (目)Phosphatocopina
- Hesslandona、Vestrogothiaなど。大顎類のステムグループ[108]もしくは甲殻類[112]。カンブリア紀に生息。
- マーレロモルフ綱 Marrellomorpha
- マーレロモルフ類[101][103]。マーレラ、ヴァコシニア、フルカ、ミメタスターなど。真節足動物[90]/大顎類[108][110]のいずれかのステムグループ。カンブリア紀 - デボン紀に生息。




- ユーシカルシノイド綱 Euthycarcinoidea
- ユーシカルシノイド類[101]。ユーシカルシヌス、ヘテロクラニア、アパンクラなど。大顎類[90]/多足類[113]のいずれかのステムグループ。カンブリア紀 - 三畳紀に生息。
-
 Euthycarcinus(旧復元)
Euthycarcinus(旧復元)

- 嚢頭綱 Thylacocephala
- 嚢頭類(ティラコセファルス類[101])。Thylacares、ドロカリス、キタカミカリス(アンキトカゾカリスの一種とされる場合もある[114])など。甲殻類としての位置は不確実[115]。オルドビス紀[116](カンブリア紀?)- 白亜紀に生息。
-

-

-

-
 Ainiktozoon(上、青)
Ainiktozoon(上、青)




- (目)Cyclida
Cyclus、Schramine、Americlusなど。甲殻類としての位置は不確実[117]。石炭紀 - 白亜紀に生息。



- モリソニア目 Mollisoniida[120]
- モリソニア類。モリソニア[121]、Thelxiope など。鋏角類/真鋏角類のいずれかのステムグループ[119][120]。カンブリア紀 - オルドビス紀に生息。

- (上綱[122])Vicissicaudata - シドネイア、エメラルデラ、ケロニエロン類、光楯類など。
- 三葉形類 Trilobitomorpha
- Petalopleura - シンダレラ、シャンダレラなど。
- Nektaspida - ナラオイアなど。
- Conciliterga - クアマイア、サペリオンなど。
- 三葉虫綱 Trilobita - 三葉虫。




旧分類
分子系統学、分岐分類学が盛行する以前には、形態に基づく以下の分類体系が使用されていた。流通している書籍と文献にもこの分類にしたがっているものも多い。よって参考・比較のため、また生物学史上の意義もあり、以下に併記する。
- 大顎亜門 Mandibulata(その後の大顎類に相当)
人間との関わり
この節の加筆が望まれています。 |
食文化
-

-

-
 販売される蜂蜜
販売される蜂蜜




節足動物は人間の食文化と深く関わっている。食材とされ、食品の生成に関わり、および食害を与えるものがある。
食材とされる節足動物の中で甲殻類は特に代表的で、カニ・エビなどの十脚類は魚介類として一般的である。それ以外の甲殻類、例えばアミ、オキアミ、フジツボなどにも食用とされる場合がある。ムカデ・クモ・サソリ、および昆虫など一般に「虫」と扱われる節足動物の中でも、地域によっては食材とされる種類がある(昆虫食)。また、特定の節足動物に頼ってできた食品もあり、蜂蜜(ミツバチによる)やミルベンケーゼ(チーズダニによる)[124]などが挙げられる。
一方で、人間の食材や食品を食害する節足動物もあり、特に農作物を食害するものは農業害虫に含まれる。このような害虫とされる種類を持つ節足動物は、バッタ(蝗害など)・カメムシ(ミナミアオカメムシなど)・アブラムシ・甲虫(コクゾウムシなど)・鱗翅類(農作物を食害するイモムシとケムシ)などの昆虫のみならず、ダニ(ハダニなど)・ヤスデ[125]にも食害を与える種類がある。
飼育



節足動物はペットや観察の対象として飼育されることが多い。その範囲は幅広く、陸生の昆虫やクモガタ類から水生の甲殻類まで及ぶ。観賞用に飼育される主な節足動物は、甲虫(カブトムシ、クワガタムシなど)・エビ(ザリガニなど)・オオツチグモなどが挙げられる。一部の分類群に対しては、それに向いている累代飼育方法や関連産品が創設され、また節足動物そのものが飼育キットとセットで販売されることもある(カブトエビ、アルテミアなど)。
医学

ヒトに対して、刺咬・吸血・接触・寄生・媒介などにより疾患を発生させ、衛生害虫に含まれる節足動物があり、以下の例が挙げられる[126]。
- 刺咬性のもの
- 吸血性のもの
- 接触性のもの
- 鱗翅類(鱗翅目/チョウ目) - ドクガやチャドクガなどの毒針毛による皮膚炎[130]。イラガなどの毒棘による皮膚炎[130]。
- 甲虫類(鞘翅目/コウチュウ目) - アオバアリガタハネカクシから分泌されるペデリンによる皮膚炎[130]。アオカミキリモドキなどから分泌されるカンタリジンによる水疱性皮膚炎[130]。ミイデラゴミムシから噴射されるガスに含まれるベンゾキノンによる皮膚炎[130]。オサムシ類やマイマイカブリから噴射されるメタアクリル酸による皮膚炎[130]。
- カメムシ類(半翅目/カメムシ目) - クサギカメムシの分泌液による皮膚炎[130]。
- サソリモドキ類(クモガタ綱サソリモドキ目) - アマミサソリモドキやタイワンサソリモドキから噴射される酢酸を含む液体による皮膚炎[130]。
脚注
注釈
- ^ 左上:三葉虫のコリハペルティス(Kolihapeltis sp.)、右上:ウミサソリのスティロヌルス(Stylonurus powriensis)、左中:サソリの1種(Scorpio maurus palmatus)、右中:カニの1種(Callinectes sapidus)、左下:ムカデの1種、右下:チョウの1種トラフアゲハ(Papilio glaucus)
- ^ 具体的には陸生を主体とする分類群(多足類、六脚類、鋏角類のクモガタ類、甲殻類の等脚類など)が中心となり、また甲殻類の中で著しく特殊化した寄生性のものは虫と呼ばれるものが多い(シタムシ、シダムシ、フクロムシ、ヒジキムシなど)。
- ^ a b c 前大脳のみを持つとされる。
- ^ 前大脳と中大脳のみを持つとされる。
出典
- ^ a b c d e f g h Daley, Allison; Antcliffe, Jonathan; Drage, Harriet; Pates, Stephen (2018-05-21). “Early fossil record of Euarthropoda and the Cambrian Explosion”. Proceedings of the National Academy of Sciences 115: 201719962. doi:10.1073/pnas.1719962115.
- ^ a b c Siebold, C. Th. E. von; Siebold, C. Th E. von; Stannius, Hermann (1848). Lehrbuch der vergleichenden Anatomie der Wirbellosen Thiere. Erster Theil. In Lehrbuch der vergleichenden Anatomie (eds C. T. von Siebold and H. Stannius). Berlin: Verlag von Veit & Comp.. pp. 679
- ^ a b Zhi-Qiang Zhang (2011), “Phylum Arthropoda von Siebold, 1848”. In: Zhang Z.-Q. (Ed.), Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, Volume 3148, Magnolia Press, Pages 99-103.
- ^ a b c d e f g h i j k l m n o p Giribet, Gonzalo; Edgecombe, Gregory D. (2019-06-17). “The Phylogeny and Evolutionary History of Arthropods”. Current Biology 29 (12): R592–R602. doi:10.1016/j.cub.2019.04.057. ISSN 0960-9822.
- ^ a b c “Arthropoda (arthropods)” (英語). Animal Diversity Web. 2018年10月24日閲覧。
- ^ a b c Giribet, Gonzalo; Edgecombe, Gregory D. (2013). Minelli, Alessandro; Boxshall, Geoffrey; Fusco, Giuseppe. eds (英語). The Arthropoda: A Phylogenetic Framework. In book: Arthropod Biology and Evolution – Molecules, Development, Morphology (pp.17-40)Chapter: The Arthropoda: a phylogenetic framework. Berlin, Heidelberg: Springer Berlin Heidelberg. pp. 17–40. doi:10.1007/978-3-642-36160-9_2. ISBN 978-3-642-36159-3
- ^ 宮崎勝己 著「節足動物における分類学の歴史」、石川良輔編、岩槻邦男・馬渡峻輔監修 編『節足動物の多様性と系統』裳華房〈バイオディバーシティ・シリーズ6〉、2008年4月11日、p.2頁。ISBN 978-4-7853-5829-7。
- ^ a b (英語) A Dictionary of Entomology. CABI. (2011). ISBN 9781845935429
- ^ a b c d e f g h Fusco, Giuseppe; Minelli, Alessandro (2013). Minelli, Alessandro; Boxshall, Geoffrey; Fusco, Giuseppe. eds (英語). Arthropod Biology and Evolution: Molecules, Development, Morphology. Berlin, Heidelberg: Springer. pp. 197–221. doi:10.1007/978-3-642-36160-9_9. ISBN 978-3-642-36160-9
- ^ a b c d e f g h i j k l m n o p q r s Javier Ortega-Hernández, Ralf Janssen, Graham E. Budd (2017-05-01). “Origin and evolution of the panarthropod head – A palaeobiological and developmental perspective” (英語). Arthropod Structure & Development 46 (3): 354–379. doi:10.1016/j.asd.2016.10.011. ISSN 1467-8039.
- ^ Khanna, D. R. (2004) (英語). Biology of Arthropoda. Discovery Publishing House. ISBN 9788171418978
- ^ Ruppert, E. E.; R. S. Fox; R. D. Barnes (2004), Invertebrate Zoology (7th ed.), Brooks/Cole, ISBN 0-03-025982-7
- ^ a b Haas, M. Susan; Brown, Susan J.; Beeman, Richard W. (2001-03-08). “Homeotic evidence for the appendicular origin of the labrum in Tribolium castaneum” (英語). Development Genes and Evolution 211 (2): 96–102. doi:10.1007/s004270000129. ISSN 0949-944X.
- ^ a b Du, Xiaoliang; Yue, Chao; Hua, Baozhen (2009-08). “Embryonic development of the scorpionfly Panorpa emarginata Cheng with special reference to external morphology (Mecoptera: Panorpidae)” (英語). Journal of Morphology 270 (8): 984–995. doi:10.1002/jmor.10736. ISSN 0362-2525.
- ^ a b c d e f g h i j k l m n o p q r s Javier, Ortega-Hernández, (2016). “Making sense of ‘lower’ and ‘upper’ stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848” (英語). Biological Reviews 91 (1): 255-273. ISSN 1464-7931.
- ^ a b c d e f g Yang, Jie; Ortega-Hernández, Javier; Legg, David A.; Lan, Tian; Hou, Jin-bo; Zhang, Xi-guang (2018-02-01). “Early Cambrian fuxianhuiids from China reveal origin of the gnathobasic protopodite in euarthropods” (英語). Nature Communications 9 (1). doi:10.1038/s41467-017-02754-z. ISSN 2041-1723.
- ^ a b Liu, Yu; Edgecombe, Gregory D.; Schmidt, Michel; Bond, Andrew D.; Melzer, Roland R.; Zhai, Dayou; Mai, Huijuan; Zhang, Maoyin et al. (2021-07-30). “Exites in Cambrian arthropods and homology of arthropod limb branches” (英語). Nature Communications 12 (1): 4619. doi:10.1038/s41467-021-24918-8. ISSN 2041-1723.
- ^ a b Boxshall, G. (2009年). “Exopodites , Epipodites and Gills in Crustaceans” (英語). www.semanticscholar.org. 2021年12月23日閲覧。
- ^ Turetzek, Natascha; Pechmann, Matthias; Schomburg, Christoph; Schneider, Julia; Prpic, Nikola-Michael (2016-01-01). “Neofunctionalization of a Duplicate dachshund Gene Underlies the Evolution of a Novel Leg Segment in Arachnids”. Molecular Biology and Evolution 33 (1): 109–121. doi:10.1093/molbev/msv200. ISSN 0737-4038.
- ^ “Appendage - an overview | ScienceDirect Topics”. www.sciencedirect.com. 2021年5月24日閲覧。
- ^ a b c Rainer F. Foelix (2011). Biology of spiders (3rd ed.). New York: Oxford University Press. ISBN 978-0-19-981324-7. OCLC 693776865
- ^ a b 村上安則 著「神経系の発生 ―ひとりでに出来上がるコンピュータ」、公益社団法人 日本動物学会 編『動物学の百科事典』丸善出版、2018年9月28日、310-311頁。ISBN 978-4621303092。
- ^ a b c d 水波誠 著「昆虫の微小脳 ―小さな脳のすごい働き」、公益社団法人 日本動物学会 編『動物学の百科事典』丸善出版、2018年9月28日、362-363頁。ISBN 978-4621303092。
- ^ Smarandache-Wellmann, Carmen Ramona (10 24, 2016). “Arthropod neurons and nervous system”. Current biology: CB 26 (20): R960–R965. doi:10.1016/j.cub.2016.07.063. ISSN 1879-0445. PMID 27780069.
- ^ a b Lamsdell, James C.; Dunlop, Jason A.. “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3): 395–418. ISSN 1467-8039.
- ^ “The tettigoniid (Orthoptera : Tettigoniidae) ear: Multiple functions and structural diversity” (英語). International Journal of Insect Morphology and Embryology 22 (2-4): 185–205. (1993-04-01). doi:10.1016/0020-7322(93)90009-P. ISSN 0020-7322.
- ^ “Acoustic Receivers: From Insect Ear to Next-Generation Sensor | ARCHIE-WeSt”. 2018年12月20日閲覧。
- ^ Yager, David; Hoy, Ron (1988-01-01). “The midline metathoracic ear of the praying mantis, Mantis religiosa”. Cell and tissue research 250: 531-541. doi:10.1007/BF00218944.
- ^ a b c Ortega-Hernández, Javier (2015-06-15). “Homology of Head Sclerites in Burgess Shale Euarthropods” (英語). Current Biology 25 (12): 1625-1631. doi:10.1016/j.cub.2015.04.034. ISSN 0960-9822.
- ^ a b Miether, Sebastian T.; Dunlop, Jason A. (2016/07). “Lateral eye evolution in the arachnids”. Arachnology 17 (2): 103-119. doi:10.13156/arac.2006.17.2.103. ISSN 2050-9928.
- ^ Kühl, Gabriele; Briggs, Derek E. G.; Rust, Jes (2009-02-06). “A Great-Appendage Arthropod with a Radial Mouth from the Lower Devonian Hunsrück Slate, Germany” (英語). Science 323 (5915): 771-773. doi:10.1126/science.1166586. ISSN 0036-8075. PMID 19197061.
- ^ Paterson, John R.; García-Bellido, Diego C.; Lee, Michael S. Y.; Brock, Glenn A.; Jago, James B.; Edgecombe, Gregory D. (2011-12). “Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes” (英語). Nature 480 (7376): 237–240. doi:10.1038/nature10689. ISSN 0028-0836.
- ^ a b c Cong, Peiyun; Ma, Xiaoya; Hou, Xianguang; Edgecombe, Gregory D.; Strausfeld, Nicholas J. (2014-07-16). “Brain structure resolves the segmental affinity of anomalocaridid appendages” (英語). Nature 513 (7519): 538–542. doi:10.1038/nature13486. ISSN 0028-0836.
- ^ Strausfeld, Nicholas J.; Ma, Xiaoya; Edgecombe, Gregory D.; Fortey, Richard A.; Land, Michael F.; Liu, Yu; Cong, Peiyun; Hou, Xianguang (2016-03). “Arthropod eyes: The early Cambrian fossil record and divergent evolution of visual systems” (英語). Arthropod Structure & Development 45 (2): 152–172. doi:10.1016/j.asd.2015.07.005.
- ^ Farrelly, C.A.; Greenaway, P. (2005-1). “The morphology and vasculature of the respiratory organs of terrestrial hermit crabs (Coenobita and Birgus): gills, branchiostegal lungs and abdominal lungs” (英語). Arthropod Structure & Development 34 (1): 63–87. doi:10.1016/j.asd.2004.11.002.
- ^ Edgecombe, Gregory D.; Giribet, Gonzalo (2007-01-01). “Evolutionary Biology of Centipedes (Myriapoda: Chilopoda)”. Annual Review of Entomology 52 (1): 151–170. doi:10.1146/annurev.ento.52.110405.091326. ISSN 0066-4170.
- ^ a b Telford, Maximilian J; Bourlat, Sarah J; Economou, Andrew; Papillon, Daniel; Rota-Stabelli, Omar (2008-04-27). “The evolution of the Ecdysozoa”. Philosophical Transactions of the Royal Society B: Biological Sciences 363 (1496): 1529–1537. doi:10.1098/rstb.2007.2243. ISSN 0962-8436. PMC 2614232. PMID 18192181.
- ^ a b Edgecombe, Gregory D. (2009-06). “Palaeontological and Molecular Evidence Linking Arthropods, Onychophorans, and other Ecdysozoa” (英語). Evolution: Education and Outreach 2 (2): 178–190. doi:10.1007/s12052-009-0118-3. ISSN 1936-6434.
- ^ a b c Giribet, Gonzalo; Edgecombe, Gregory D. (2017-09-01). “Current Understanding of Ecdysozoa and its Internal Phylogenetic Relationships”. Integrative and Comparative Biology 57 (3): 455–466. doi:10.1093/icb/icx072. ISSN 1540-7063.
- ^ Haeckel E. 1866. Generelle Morphologie der Organismen. Allgemeine Grundzüge der Organischen formen-wissenschaft, mechanisch begründet durch die von Charles Darwin reformirte descendenztheorie, 2 vols. Berlin: Georg Reimer.
- ^ Snodgrass, Robert Evans. 1938. "Evolution of the annelida onychophora and arthropoda." Smithsonian Miscellaneous Collections. 97 (6):1–159.
- ^ Boudreaux, H. Bruce (1979). Arthropod phylogeny with special reference to insects. Internet Archive. New York : Wiley. ISBN 978-0-471-04290-7
- ^ Dzik, Jerzy; Krumbiegel, Günter (1989). “The oldest ‘onychophoran’Xenusion: a link connecting phyla?” (英語). Lethaia 22 (2): 169–181. doi:10.1111/j.1502-3931.1989.tb01679.x. ISSN 1502-3931.
- ^ MONGE-NAJERA, JULIAN (1995-05-01). “Phylogeny, biogeography and reproductive trends in the Onychophora”. Zoological Journal of the Linnean Society 114 (1): 21–60. doi:10.1111/j.1096-3642.1995.tb00111.x. ISSN 0024-4082.
- ^ LANKESTER, E. RAY (1904-03-01). “Memoirs: The Structure and Classification of the Arthropoda”. Journal of Cell Science s2-47 (188): 523–582. doi:10.1242/jcs.s2-47.188.523. ISSN 0021-9533.
- ^ Vandel A. 1949. Arthropodes: Généralités, composition de l’embranchement. In: Grassé pp, ed. Traité de Zoologie, 6. Paris: Masson, 79–158.
- ^ SALVADOR V. PERIS 1971. Los órdenes de los artrópodos. Trab. Cátedra Artrópodos (Fac. Biología, Univ. Complutense) 1. 1-121
- ^ Aguinaldo, Anna Marie A.; Turbeville, James M.; Linford, Lawrence S.; Rivera, Maria C.; Garey, James R.; Raff, Rudolf A.; Lake, James A. (1997-05). “Evidence for a clade of nematodes, arthropods and other moulting animals” (英語). Nature 387 (6632): 489–493. doi:10.1038/387489a0. ISSN 0028-0836.
- ^ Adoutte, A; Balavoine, Guillaume; Lartillot, N; Lespinet, O; Prud'homme, B; de Rosa, R (2000-05-01). The new animal phylogeny: Reliability and implications. 97
- ^ Seaver, Elaine C.; Kaneshige, Lori M. (2006-01-01). “Expression of ‘segmentation’ genes during larval and juvenile development in the polychaetes Capitella sp. I and H. elegans” (英語). Developmental Biology 289 (1): 179–194. doi:10.1016/j.ydbio.2005.10.025. ISSN 0012-1606.
- ^ Mayer, Georg (2006-03-01). “Origin and differentiation of nephridia in the Onychophora provide no support for the Articulata” (英語). Zoomorphology 125 (1): 1–12. doi:10.1007/s00435-005-0006-5. ISSN 1432-234X.
- ^ Chipman, Ariel D. (2010). “Parallel evolution of segmentation by co-option of ancestral gene regulatory networks” (英語). BioEssays 32 (1): 60–70. doi:10.1002/bies.200900130. ISSN 1521-1878.
- ^ Storch, Volker; Jamieson, Barrie G. M. (1992-02-01). “Further spermatological evidence for including the pentastomida (tongue worms) in the Crustacea” (英語). International Journal for Parasitology 22 (1): 95–108. doi:10.1016/0020-7519(92)90085-Y. ISSN 0020-7519.
- ^ Lavrov, Dennis V.; Brown, Wesley M.; Boore, Jeffrey L. (2004-03-07). “Phylogenetic position of the Pentastomida and (pan)crustacean relationships”. Proceedings of the Royal Society of London. Series B: Biological Sciences 271 (1538): 537–544. doi:10.1098/rspb.2003.2631. PMC PMC1691615. PMID 15129965.
- ^ Nielsen, C. (1995). Animal Evolution, Interrelationships of the Living Phyla. Oxford University Press, Oxford.
- ^ Persson, Dennis K.; Halberg, Kenneth A.; Jørgensen, Aslak; Møbjerg, Nadja; Kristensen, Reinhardt M. (2012-11-01). “Neuroanatomy of Halobiotus crispae (Eutardigrada: Hypsibiidae): Tardigrade brain structure supports the clade panarthropoda” (英語). Journal of Morphology 273 (11). doi:10.1002/jmor.20054/abstract. ISSN 1097-4687.
- ^ Hyun Ryu, Shi; Lee, Jimin; Jang, Kuem-Hee; Hwa Choi, Eun; Ju Park, Shin; Chang, Cheon; Kim, Won; Hwang, Ui Wook (2008-01-01). “Partial mitochondrial gene arrangements support a close relationship between Tardigrada and Arthropoda”. Molecules and cells 24: 351-357.
- ^ Omar Rota-Stabelli, Ehsan Kayal, Dianne Gleeson, Jennifer Daub, Jeffrey L. Boore, Maximilian J. Telford, Davide Pisani, Mark Blaxter & Dennis V. Lavrov (2010). “Ecdysozoan mitogenomics: evidence for a common origin of the legged invertebrates, the Panarthropoda”. Genome Biology and Evolution 2: 425–440. doi:10.1093/gbe/evq030. PMID 20624745.
- ^ Ahlrichs, Wilko (1995) (German). Ultrastruktur und Phylogenie von Seison nebaliae (Grube 1859) und Seison annulatus (Claus 1876) : Hypothesen zu phylogenetischen Verwandtschaftsverhältnissen innerhalb der Bilateria. Cuvillier. ISBN 978-3-89588-295-1. OCLC 40302795
- ^ HejnolAndreas 著、WanningerAndreas 編(英語)『Cycloneuralia』Springer、2015年、1–13頁。doi:10.1007/978-3-7091-1865-8_1。ISBN 978-3-7091-1865-8。2021年12月23日閲覧。
- ^ “Segmentation in Tardigrada and diversification of segmental patterns in Panarthropoda” (英語). Arthropod Structure & Development 46 (3): 328–340. (2017-05-01). doi:10.1016/j.asd.2016.10.005. ISSN 1467-8039.
- ^ a b c d e f g h i Zeng, Han; Zhao, Fangchen; Niu, Kecheng; Zhu, Maoyan; Huang, Diying (2020-12). “An early Cambrian euarthropod with radiodont-like raptorial appendages” (英語). Nature 588 (7836): 101–105. doi:10.1038/s41586-020-2883-7. ISSN 1476-4687.
- ^ a b c Budd, Graham (1993-08). “A Cambrian gilled lobopod from Greenland” (英語). Nature 364 (6439): 709–711. doi:10.1038/364709a0. ISSN 0028-0836.
- ^ BUDD, GRAHAM E. (1996-03). “The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group” (英語). Lethaia 29 (1): 1–14. doi:10.1111/j.1502-3931.1996.tb01831.x. ISSN 0024-1164.
- ^ a b Van Roy, Peter; Daley, Allison C.; Briggs, Derek E. G. (2015-03-11). “Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps” (英語). Nature 522 (7554): 77–80. doi:10.1038/nature14256. ISSN 0028-0836.
- ^ a b c d e Edgecombe, Gregory D. (2020-11-02). “Arthropod Origins: Integrating Paleontological and Molecular Evidence”. Annual Review of Ecology, Evolution, and Systematics 51 (1): 1–25. doi:10.1146/annurev-ecolsys-011720-124437. ISSN 1543-592X.
- ^ Collins, Desmond (1996/03). “The “evolution” of Anomalocaris and its classification in the arthropod class Dinocarida (nov.) and order Radiodonta (nov.)” (英語). Journal of Paleontology 70 (2): 280–293. doi:10.1017/S0022336000023362. ISSN 0022-3360.
- ^ Aria, Cédric; Caron, Jean-Bernard (2015). “Cephalic and limb anatomy of a new Isoxyid from the Burgess Shale and the role of "stem bivalved arthropods" in the disparity of the frontalmost appendage”. PloS One 10 (6): e0124979. doi:10.1371/journal.pone.0124979. ISSN 1932-6203. PMC 4454494. PMID 26038846.
- ^ Vannier, Jean; Liu, Jianni; Lerosey-Aubril, Rudy; Vinther, Jakob; Daley, Allison C. (2014-05-02). “Sophisticated digestive systems in early arthropods” (英語). Nature Communications 5 (1): 1–9. doi:10.1038/ncomms4641. ISSN 2041-1723.
- ^ a b Størmer, Leif (1944) (English). On the relationships and phylogeny of fossil and recent Arachnomorpha: a comparative study on Arachnida, Xiphosura, Eurypterida, Trilobita, and other fossil Arthropoda. Oslo: Jacob Dybwad. OCLC 961296639
- ^ Kraus, O. (1998). Fortey, R. A.; Thomas, R. H.. eds (英語). Arthropod Relationships. Dordrecht: Springer Netherlands. pp. 295–303. doi:10.1007/978-94-011-4904-4_22. ISBN 978-94-011-4904-4
- ^ Telford, Maximilian J.; Thomas, Richard H. (1998-09-01). “Expression of homeobox genes shows chelicerate arthropods retain their deutocerebral segment” (英語). Proceedings of the National Academy of Sciences 95 (18): 10671–10675. ISSN 0027-8424. PMID 9724762.
- ^ Mittmann, Beate; Scholtz, Gerhard (2003-02-01). “Development of the nervous system in the "head" of Limulus polyphemus (Chelicerata: Xiphosura): morphological evidence for a correspondence between the segments of the chelicerae and of the (first) antennae of Mandibulata” (英語). Development Genes and Evolution 213 (1): 9?17. doi:10.1007/s00427-002-0285-5. ISSN 1432-041X.
- ^ “Immunohistochemical localization of neurotransmitters in the nervous system of larval Limulus polyphemus (Chelicerata, Xiphosura): evidence for a conserved protocerebral architecture in Euarthropoda” (英語). Arthropod Structure & Development 34 (3): 327–342. (2005-07-01). doi:10.1016/j.asd.2005.01.006. ISSN 1467-8039.
- ^ Sharma, Prashant P.; Tarazona, Oscar A.; Lopez, Davys H.; Schwager, Evelyn E.; Cohn, Martin J.; Wheeler, Ward C.; Extavour, Cassandra G. (2015-06-07). “A conserved genetic mechanism specifies deutocerebral appendage identity in insects and arachnids”. Proceedings of the Royal Society B: Biological Sciences 282 (1808): 20150698. doi:10.1098/rspb.2015.0698. PMC PMC4455815. PMID 25948691.
- ^ Giribet, Gonzalo; Ribera, Carles (2000-06). “A Review of Arthropod Phylogeny: New Data Based on Ribosomal DNA Sequences and Direct Character Optimization” (英語). Cladistics 16 (2): 204–231. doi:10.1111/j.1096-0031.2000.tb00353.x. ISSN 0748-3007.
- ^ Nardi, Francesco; Spinsanti, Giacomo; Boore, Jeffrey L.; Carapelli, Antonio; Dallai, Romano; Frati, Francesco (2003-03-21). “Hexapod Origins: Monophyletic or Paraphyletic?” (英語). Science 299 (5614): 1887–1889. doi:10.1126/science.1078607. ISSN 0036-8075. PMID 12649480.
- ^ Regier, Jerome C.; Shultz, Jeffrey W.; Kambic, Robert E. (2005-02-22). “Pancrustacean phylogeny: hexapods are terrestrial crustaceans and maxillopods are not monophyletic”. Proceedings of the Royal Society B: Biological Sciences 272 (1561): 395–401. doi:10.1098/rspb.2004.2917. ISSN 0962-8452. PMC 1634985. PMID 15734694.
- ^ a b Regier, Jerome C.; Shultz, Jeffrey W.; Zwick, Andreas; Hussey, April; Ball, Bernard; Wetzer, Regina; Martin, Joel W.; Cunningham, Clifford W. (2010-02). “Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences” (英語). Nature 463 (7284): 1079–1083. doi:10.1038/nature08742. ISSN 0028-0836.
- ^ Oakley, Todd H.; Wolfe, Joanna M.; Lindgren, Annie R.; Zaharoff, Alexander K. (2012-09-12). “Phylotranscriptomics to Bring the Understudied into the Fold: Monophyletic Ostracoda, Fossil Placement, and Pancrustacean Phylogeny” (英語). Molecular Biology and Evolution 30 (1): 215–233. doi:10.1093/molbev/mss216. ISSN 1537-1719.
- ^ Harrison, Jon Fewell (2015年). “Linking Insects with Crustacea : Comparative Physiology of the Pancrustacea Organized by” (英語). 2018年10月24日閲覧。
- ^ Lozano-Fernandez, Jesus; Giacomelli, Mattia; Fleming, James F; Chen, Albert; Vinther, Jakob; Thomsen, Philip Francis; Glenner, Henrik; Palero, Ferran et al. (2019-08-01). “Pancrustacean Evolution Illuminated by Taxon-Rich Genomic-Scale Data Sets with an Expanded Remipede Sampling”. Genome Biology and Evolution 11 (8): 2055–2070. doi:10.1093/gbe/evz097. ISSN 1759-6653.
- ^ Dunlop, Jason; Borner, Janus; Burmester, Thorsten (2014-02-27) (英語). 16 Phylogeny of the Chelicerates: Morphological and molecular evidence. De Gruyter. doi:10.1515/9783110277524.399. ISBN 978-3-11-027752-4
- ^ Hassanin, Alexandre; Prieur, Stéphanie le; Bonillo, Céline; Krapp, Franz; Corbari, Laure; Sabroux, Romain (2017-02-24). “Biodiversity and phylogeny of Ammotheidae (Arthropoda: Pycnogonida)” (英語). European Journal of Taxonomy 0 (286). doi:10.5852/ejt.2017.286. ISSN 2118-9773.
- ^ Giribet, Gonzalo; Edgecombe, Gregory D.; Wheeler, Ward C. (2001-09). “Arthropod phylogeny based on eight molecular loci and morphology” (英語). Nature 413 (6852): 157–161. doi:10.1038/35093097. ISSN 1476-4687.
- ^ Dunlop, J. A.; Arango, C. P. (2005). “Pycnogonid affinities: A review”. Journal of Zoological Systematics and Evolutionary Research 43: 8–21. doi:10.1111/j.1439-0469.2004.00284.x.
- ^ Lamsdell, James (2013-01-01). “Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura”. Zoological Journal of the Linnean Society 167: 1–27. doi:10.1111/j.1096-3642.2012.00874.x.
- ^ “(PDF) Heads, Hox and the phylogenetic position of trilobites. Crustacea and Arthropod Phylogeny. –” (英語). ResearchGate. 2018年10月24日閲覧。
- ^ a b Scholtz, Gerhard; Staude, Andreas; Dunlop, Jason A. (2019-06-07). “Trilobite compound eyes with crystalline cones and rhabdoms show mandibulate affinities” (英語). Nature Communications 10 (1): 1-7. doi:10.1038/s41467-019-10459-8. ISSN 2041-1723.
- ^ a b c d e f g Aria, Cédric; Caron, Jean-Bernard (2017-05). “Burgess Shale fossils illustrate the origin of the mandibulate body plan” (英語). Nature 545 (7652): 89–92. doi:10.1038/nature22080. ISSN 1476-4687.
- ^ a b Tanaka, Gengo; Hou, Xianguang; Ma, Xiaoya; Edgecombe, Gregory D.; Strausfeld, Nicholas J. (2013-10). “Chelicerate neural ground pattern in a Cambrian great appendage arthropod” (英語). Nature 502 (7471): 364–367. doi:10.1038/nature12520. ISSN 0028-0836.
- ^ Legg, David (2013-06). The impact of fossils on arthropod phylogeny.
- ^ 宮崎勝己「節足動物全体の分類体系・系統の現状」石川良輔編『節足動物の多様性と系統』〈バイオディバーシティ・シリーズ〉第6巻、岩槻邦男・馬渡峻輔監修、裳華房、2008年、11-27頁。
- ^ a b c d Hou, Xianguang; Bergström, Jan (1997). Arthropods of the Lower Cambrian Chengjiang fauna, southwest China. Oslo: Scandinavian University Press. ISBN 82-00-37693-1. OCLC 38305908
- ^ a b 鈴木雄太郎「三葉虫綱」石川良輔編『節足動物の多様性と系統』〈バイオディバーシティ・シリーズ〉第6巻、岩槻邦男・馬渡峻輔監修、裳華房、2008年、114-121頁。
- ^ a b c d e f 大塚攻、田中隼人「顎脚類(甲殻類)の分類と系統に関する研究の最近の動向」『タクサ:日本動物分類学会誌』第48巻、日本動物分類学会、2020年、49–62頁、doi:10.19004/taxa.48.0_49。
- ^ Basset, Yves; Cizek, Lukas; Cuenoud, Philippe; Didham, Raphael; Guilhaumon, Francois; Missa, Olivier; Novotny, Vojtech; Ødegaard, Frode et al. (2012-12-14). “Arthropod Diversity in a Tropical Forest”. Science 338: 1481–1484. doi:10.1126/science.1226727.
- ^ “What are trilobites? - Australian Museum” (英語). australianmuseum.net.au. 2018年10月25日閲覧。
- ^ a b 金子, 隆一 (2012) (Japanese). ぞわぞわした生きものたち: 古生代の巨大節足動物. 東京: ソフトバンククリエイティブ. ISBN 978-4-7973-4411-0. OCLC 816905375
- ^ a b “岡山県野生生物目録2019 - 岡山県ホームページ(自然環境課)”. www.pref.okayama.jp. 2019年8月16日閲覧。
- ^ a b c d e 加藤太一 (2017) (Japanese). 古生物. 東京: 学研プラス. ISBN 978-4-05-204576-9. OCLC 992701133
- ^ 土屋, 健; かわさき, しゅんいち; 田中, 源吾 (2020) (Japanese). アノマロカリス解体新書. ISBN 978-4-89308-928-1. OCLC 1141813539
- ^ a b 群馬県立自然史博物館 (2020) (Japanese). 大むかしの生きもの. 東京: 講談社. ISBN 978-4-06-518985-6. OCLC 1163637303
- ^ 土屋, 健; 土屋, 香; 芝原, 暁彦 (2021) (Japanese). ゼロから楽しむ古生物: 姿かたちの移り変わり. ISBN 978-4-297-12228-7. OCLC 1262176890
- ^ 小林快次 (2014) (Japanese). 大昔の生きもの. 東京: ポプラ社. ISBN 978-4-591-14071-0. OCLC 883613863
- ^ 土屋健; 群馬県立自然史博物館 (2017). 生命史図譜 = History of life. ISBN 978-4-7741-9075-4. OCLC 995843160
- ^ a b Legg, David A.; Sutton, Mark D.; Edgecombe, Gregory D.; Caron, Jean-Bernard (2012-12-07). “Cambrian bivalved arthropod reveals origin of arthrodization”. Proceedings of the Royal Society B: Biological Sciences 279 (1748): 4699–4704. doi:10.1098/rspb.2012.1958. PMC PMC3497099. PMID 23055069.
- ^ a b c d e f Legg, David A.; Sutton, Mark D.; Edgecombe, Gregory D. (2013). “Arthropod fossil data increase congruence of morphological and molecular phylogenies” (英語). Nature Communications 4 (1). ISSN 2041-1723.
- ^ a b Vannier, Jean; Aria, Cédric; Taylor, Rod S.; Caron, Jean-Bernard (2018). “Waptia fieldensis Walcott, a mandibulate arthropod from the middle Cambrian Burgess Shale”. Royal Society Open Science 5 (6): 172206. doi:10.1098/rsos.172206. PMC PMC6030330. PMID 30110460.
- ^ a b Zhai, Dayou; Williams, Mark; Siveter, David J.; Harvey, Thomas H. P.; Sansom, Robert S.; Gabbott, Sarah E.; Siveter, Derek J.; Ma, Xiaoya et al. (2019-09-03). “Variation in appendages in early Cambrian bradoriids reveals a wide range of body plans in stem-euarthropods” (英語). Communications Biology 2 (1): 1–6. doi:10.1038/s42003-019-0573-5. ISSN 2399-3642.
- ^ Hou, Xianguang; Williams, Mark; Siveter, David J.; Siveter, Derek J.; Aldridge, Richard J.; Sansom, Robert S. (2010-06-22). “Soft-part anatomy of the Early Cambrian bivalved arthropods Kunyangella and Kunmingella: significance for the phylogenetic relationships of Bradoriida”. Proceedings of the Royal Society B: Biological Sciences 277 (1689): 1835-1841. doi:10.1098/rspb.2009.2194. PMC PMC2871875. PMID 20181565.
- ^ Fossils and Strata, Morphology, Ontogeny and Phylogeny of the Phosphatocopina (Crustacea) from the Upper Cambrian Orsten of Sweden.. Waloszek, Dieter., Maas, Andreas., Muller, Klaus.. Wiley. (2009). ISBN 1-282-11829-3. OCLC 741343594
- ^ Edgecombe, Gregory D.; Strullu-Derrien, Christine; Góral, Tomasz; Hetherington, Alexander J.; Thompson, Christine; Koch, Markus (2020-04-01). “Aquatic stem group myriapods close a gap between molecular divergence dates and the terrestrial fossil record” (英語). Proceedings of the National Academy of Sciences. doi:10.1073/pnas.1920733117. ISSN 0027-8424. PMID 32253305.
- ^ Laville, Thomas; Smith, Christopher P. A.; Forel, Marie-Béatrice; Brayard, Arnaud; Charbonnier, Sylvain (2021-02-25). “REVIEW OF EARLY TRIASSIC THYLACOCEPHALA” (英語). RIVISTA ITALIANA DI PALEONTOLOGIA E STRATIGRAFIA 127 (1). doi:10.13130/2039-4942/15188. ISSN 2039-4942.
- ^ Lange, Sven; Schram, Frederick R.; Steeman, Fedor A.; Hof, Cees H. J. (2001). “New Genus and Species from the Cretaceous of Lebanon Links the Thylacocephala To the Crustacea” (英語). Palaeontology 44 (5): 905–912. doi:10.1111/1475-4983.00207. ISSN 1475-4983.
- ^ Roy, Peter Van; Rak, Štěpán; Budil, Petr; Fatka, Oldřich (2021). “Upper Ordovician Thylacocephala (Euarthropoda, Eucrustacea) from Bohemia indicate early ecological differentiation” (英語). Papers in Palaeontology 7 (3): 1727–1751. doi:10.1002/spp2.1363. ISSN 2056-2802.
- ^ Schädel, Mario; Haug, Joachim T. (2020/05). “A new interpretation of the enigmatic fossil arthropod Anhelkocephalon handlirschi Bill, 1914 – important insights in the morphology of Cyclida”. Palaeodiversity 13 (1): 69–81. doi:10.18476/pale.v13.a7. ISSN 1867-6294.
- ^ a b Aria, Cédric; Caron, Jean-Bernard (2017-12). “Mandibulate convergence in an armoured Cambrian stem chelicerate” (英語). BMC Evolutionary Biology 17 (1): 261. doi:10.1186/s12862-017-1088-7. ISSN 1471-2148. PMC PMC5738823. PMID 29262772.
- ^ a b Aria, Cédric; Caron, Jean-Bernard (2019-09). “A middle Cambrian arthropod with chelicerae and proto-book gills” (英語). Nature 573 (7775): 586–589. doi:10.1038/s41586-019-1525-4. ISSN 1476-4687.
- ^ a b Lerosey-Aubril, Rudy; Skabelund, Jacob; Ortega-Hernández, Javier (2020-04-09). “Revision of the mollisoniid chelicerate(?) Thelxiope , with a new species from the middle Cambrian Wheeler Formation of Utah” (英語). PeerJ 8: e8879. doi:10.7717/peerj.8879. ISSN 2167-8359.
- ^ a b Gould, Stephen Jay; 渡辺, 政隆 (2000) (Japanese). ワンダフル・ライフ: バージェス頁岩と生物進化の物語. ISBN 978-4-15-050236-2. OCLC 676428996
- ^ a b Lerosey-Aubril, Rudy; Zhu, Xuejian; Ortega-Hernández, Javier (2017-12). “The Vicissicaudata revisited – insights from a new aglaspidid arthropod with caudal appendages from the Furongian of China” (英語). Scientific Reports 7 (1): 11117. doi:10.1038/s41598-017-11610-5. ISSN 2045-2322. PMC PMC5593897. PMID 28894246.
- ^ Ortega-Hernández, Javier; Azizi, Abdelfattah; Hearing, Thomas W.; Harvey, Thomas H. P.; Edgecombe, Gregory D.; Hafid, Ahmid; El Hariri, Khadija (2017-02-17). “A xandarellid artiopodan from Morocco – a middle Cambrian link between soft-bodied euarthropod communities in North Africa and South China” (英語). Scientific Reports 7 (1). doi:10.1038/srep42616. ISSN 2045-2322.
- ^ “Manufaktur” (ドイツ語). Würchwitzer Milbenkäse: Der lebendigste Käse der Welt.. 2019年8月17日閲覧。
- ^ Ebregt, E.; Struik, P. C.; Odongo, B.; Abidin, P. E. (2005-01-01). “Pest damage in sweet potato, groundnut and maize in north-eastern Uganda with special reference to damage by millipedes (Diplopoda)”. NJAS - Wageningen Journal of Life Sciences 53 (1): 49–69. doi:10.1016/S1573-5214(05)80010-7. ISSN 1573-5214.
- ^ 夏秋優『Dr.夏秋の臨床図鑑 虫と皮膚炎』学研プラス、2013年、10頁。
- ^ a b c d e 夏秋優『Dr.夏秋の臨床図鑑 虫と皮膚炎』学研プラス、2013年、13頁。
- ^ a b c d 夏秋優『Dr.夏秋の臨床図鑑 虫と皮膚炎』学研プラス、2013年、14頁。
- ^ “CDC - DPDx - American Trypanosomiasis” (英語). www.cdc.gov (2017年12月19日). 2019年1月26日閲覧。
- ^ a b c d e f g h 夏秋優『Dr.夏秋の臨床図鑑 虫と皮膚炎』学研プラス、2013年、15頁。
参考文献
- 石川良輔編、岩槻邦男・馬渡峻輔監修 『節足動物の多様性と系統』 裳華房、2008年、495頁、ISBN 978-4-7853-5829-7
- 白山義久編 『無脊椎動物の多様性と系統(節足動物を除く)』 裳華房、2002年、ISBN 4-7853-5828-9