
一次聴覚野
| 脳: 一次聴覚野 | |
|---|---|
 ヒトの脳の、ブロードマンの脳地図における41野と42野。一次聴覚皮質にあたる。  | |
| 名称 | |
| 日本語 | 一次聴覚野 |
| 英語 | Primary auditory cortex |
| 略号 | AI, A1 |
| 関連情報 | |
| Brede Database | 階層関係、座標情報 |
| NeuroNames | 関連情報一覧 |
一次聴覚野(いちじちょうかくや、英: Primary auditory cortex)は脳の領域の1つで、聴覚 (音) 情報の処理を担う領域である。 側頭葉の上側頭回、横側頭回に位置し、視床の内側膝状体から聴放線を通じて、信号を受けている。
機能
[編集]他の一次感覚皮質領域と同様に、音の感覚はそれが大脳皮質領域に伝わり、処理されて初めて知覚される。この根拠として、脳腫瘍や脳梗塞によって皮質領域に障害を負ってしまったと見られる患者の研究や、冷却や局所的な薬物投与によって皮質領域を非活性化する手法を用いた動物実験が存在する。一次聴覚野に損傷を受けた患者は音に対するアウェアネスを失っているが、聴覚脳幹と中脳による皮質下の処理が大部分を占める、音に対する反射的反応の能力は残されている。
聴覚野にあるニューロンは、それぞれの感受性が最も高い周波数に従って構造を成している。聴覚野の一端にあるニューロンは最も低い周波数に反応し、もう一端のニューロンは最も高い周波数に反応する。また、視覚野のように、聴覚野には異なる複数の領域が存在している。それぞれの領域は解剖学的に異なり、それぞれが完全な"周波数のマップ"を持っている。この周波数のマップ (トノトピー・マップ) の目的はよく分かっておらず、聴覚系の感覚上皮 (蝸牛) はその周波数に従って並んでいるという点を反映していると考えられている。聴覚野は聴覚的な"対象"の同定や分離、空間における音源の位置の同定などに関わっている。
ヒト脳機能イメージングによって、この脳領域の周辺部が音高を同定しようとする際に活性化することが分かっている。それぞれの細胞は特定の周波数、またはその周波数の倍数の音(倍音)に対して興奮する。
一次聴覚野はブロードマンの脳地図における41野と42野とおおよそ同一である。この領域は上側頭回の後半分と、外側溝の奥にある横側頭回 (またはヘッシェル回 (Heschl's gyrus))と呼ばれる領域である。
一次聴覚野は側頭葉に位置している。ヒトの大脳皮質には、頭頂葉の前部など他にも音の処理に関わる領域が存在する。動物実験によって、大脳皮質の聴覚野は視床の聴覚領域から上ってくる入力情報を受け取っていることや、この経路が同側と反体側で相互接続されていることが分かっている。また、聴覚野は構造的にも機能的にも異なる領域から成り立っている[1]。
聴覚野の領域の数は種によって異なっている。ネズミ目では最近まで数領域しか分かっていなかったが現在はマウスやラットを中心に6〜7領域が同定されている[2]。アカゲザルでは15領域にもなる。現時点ではヒトの聴覚野の領域の数、位置、構造はまだよく分かっていない。ヒトの聴覚野について知られていることは、霊長類を含む哺乳類の電気生理学的研究からの推測とヒトの脳機能イメージングが基礎になっている。
オーケストラやジャズバンドのそれぞれの楽器が同じ音符を演奏した時、それぞれの音の質は異なっているにもかかわらず、演奏家はそれぞれの音符の音高が同一であると認識出来る。聴覚野のニューロンは音高に対して反応することが可能である。マーモセットを用いた研究では、音高特異的なニューロンが一次聴覚野の上外側端付近に存在することが分かっている。この音高特異的な領域はヒトの脳機能イメージング研究でも見つかっている[3][4]。
聴覚野は低次領域や耳からの入力を受けるだけでなく、出力も行っている。
ブロードマンの脳地図における41野
[編集]この領域は前横側頭領域 (anterior transverse temporal area 41)とも呼ばれる。細胞構築学的には、大脳皮質の側頭部の下位領域で、側頭葉の背側面にある外側溝壁の前横側頭回に存在する。41野は内側をブロードマンの脳地図における52野に接し、外側を42野に接している。(Brodmann-1909)
ブロードマンの脳地図における42野
[編集]この領域は後横側頭領域 (posterior transverse temporal area 42)とも呼ばれる。細胞構築学的には、大脳皮質の側頭部の下位領域で、側頭葉の背側面にある外側溝壁に位置する。42野は内側を41野に接し、外側を上側頭領域 (22野) に接する。(Brodmann-1909)
聴覚系との関連
[編集]
聴覚野は脳において最も強く組織化された音処理のユニットである。この皮質領域は聴覚、及びヒトにおける言語、音楽の処理の神経的な要所となっている。
聴覚野は一次、二次、三次聴覚野の3領域に分けられる。これらの領域は同心円状に並び、一次聴覚野が内側、三次聴覚野が外側に存在する。
一次聴覚野はトノトピーに従って構造を成している。つまり、聴覚野のある領域のニューロンは特定の周波数の音に対して反応する。このことは聴覚系の経路を通じて保存されている興味深い機能である。この領域は音高や音量などの、音楽の基本的な部分を同定することが出来ると考えられる。このことは視床の内側膝状体からの直接の入力を受けていることからも分かる。二次聴覚野はハーモニー、メロディ、リズムのパターンの処理を担っていることが示唆されている。三次聴覚野は全てを音楽の全体的な体験へと統合する役割を担うと推測されている[5]。
クリンケ (Klinke) らによる先天的に聾唖の子猫の誘起電位の研究では、電場電位を利用して聴覚野の皮質可塑性が計測された。先天的に聾唖の子猫に人工内耳と音処理装置を埋め込んで刺激した結果、この子猫から計測される電場電位は正常な聴覚の猫よりも強いものとなった[6]。この結果は、音楽教育を受けた生徒は受けてない生徒に比べて皮質の活動が増加するとしたエッカート (Eckart Altenmuller) らによる研究と一致する[7]。
聴覚野はガンマ波周波数に関して不思議な現象を示す。3、4周期の40ヘルツのクリック音に曝された被験者の脳波には、他の刺激では現れない異常なスパイクが現れる。この周波数に関連して生じる神経活動のスパイクは、聴覚野のトノトピーに依存するものではない。この周波数は脳の特定領域の“共鳴周波数”であると考えられ、この活動は視覚野にも影響を及ぼしている[8]。
ガンマ波 (20 から 40 Hz の脳波) は感覚的な出来事の知覚や、認識の処理の際に現れる。ニーフ (Knief) らの2000年の研究では、ヤンキードゥードゥル (Yankee Doodle : アルプス一万尺) やフレール・ジャック (Frere Jacques : かねがなる) といったよく知られている曲から8音を、まず被験者に呈示した。その次に、ランダムに6、7番目の音符を省略して呈示し、その時の脳活動を脳波計と脳磁図を用いて調べた。その結果、聴覚課題によって発生するガンマ波は、被験者のこめかみ付近で観測された。省略された刺激に対する反応 (OSR : omitted stimulus response) は、わずかに異なる位置で計測され、その位置は通常の曲に比べて、7 mm前方、13 mm内側、13 mm上方であった。また、計測されるガンマ波は通常の曲に比べ弱いものであった。省略された6、または7番目の音に対する反応の変化は、特に右大脳半球で大きかった[9]。右側の一次聴覚野はより調性音楽に対する感受性が高いとされる一方、左側の一次聴覚野は特に音声言語の音のわずかな順番の違いにより感受性が高いとされている。
幻聴はガンマ周波数帯に類似した (完全に同一ではない) オシレーションを引き起こす。スパーリング (Sperling) は自身の2004年の研究により、幻聴が12.5から30 Hz の脳波を引き起こすことを示した。この周波数帯は統合失調症患者の右側一次聴覚野の活動を、13人の対照群と比較した時に観測された。この実験は、被験者に頭の中で曲を思い出してもらい、実際には音を聞いていないにもかかわらず、メロディやリズムや音全体の体験を経験してもらう過去の実験と対になるものである。統合失調症患者は幻聴を感じている時、一次聴覚野が活性化していた。このことは、曲を思い出してもらう際は三次聴覚野がわずかに活動することと比べて対照的な結果となった [10]。 このことから、一次聴覚野の人工的な刺激は非常にリアルな幻聴を引き起こすと考えられる。また、全ての音と音楽の情報が三次聴覚野へと終結することで、聴覚情報の魅力的な結びつきを生み出していると考えられている。この理論が正しいとすれば、三次聴覚野に損傷を受けた人や、この領域の機能を人工的に低下させた人の研究をすることは興味深いものとなるだろう。しかし三次聴覚野は他の低次の聴覚野を取り囲むように輪状に存在するため、このような実験は非常に難しいものとなる。
音色は聴覚野の他にも多くの領域で知覚される。その中でも特に興味深い領域として前頭前皮質内側吻側部 (rostromedial prefrontal cortex) がある[11]。ジャナタ (Janata) らは自身の2002年の研究において、fMRIを用いて音色の処理をする際に活動する脳の領域を調べた。その結果、通常は聴覚処理と関連がないと考えられていたいくつかの領域でも活動の変化が見られた。前頭前皮質内側吻側部は内側前頭前皮質の下位領域で、扁桃体へと投射し、負の感情の抑制を助けていると考えられている[12]。内側前頭前皮質は衝動的な10代と温和な大人を比較した際、大きく発達の異なる領域であると考えられている。前頭前皮質内側吻側部の音色に対する感受性は、この領域が共鳴する音や音楽の周波数や音色に対して活性化することを意味する。このことは音楽によって人々の魂 (見方によっては大脳辺縁系) が安らぎを得るメカニズムとして考えられるかもしれない。
画像
[編集]-
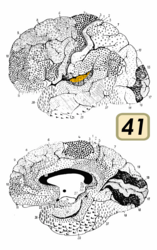 ブロードマンの脳地図における41野。聴覚野にあたる。
ブロードマンの脳地図における41野。聴覚野にあたる。 -
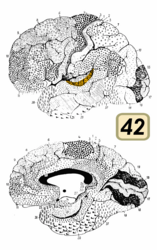 ブロードマンの脳地図における42野。聴覚野にあたる。
ブロードマンの脳地図における42野。聴覚野にあたる。 -
 ブロードマンの脳地図における22野。22野の後部(図中オレンジ色の部分の右半分ほど)はウェルニッケ野(聴覚性言語野)にあたる。
ブロードマンの脳地図における22野。22野の後部(図中オレンジ色の部分の右半分ほど)はウェルニッケ野(聴覚性言語野)にあたる。 -





関連項目
[編集]参考文献
[編集]- ^ Cant, NB (Jun 15, 2003). “Parallel auditory pathways: projection patterns of the different neuronal populations in the dorsal and ventral cochlear nuclei”. Brain Res Bull. 60 (5-6): 457-74. doi:10.1016/S0361-9230(03)00050-9.
- ^ Tsukano, H., Horie, M., Ohga, S., Takahashi, K., Kubota, Y., Hishida, R., Takebayashi, H., Shibuki, K. (2017) "Reconsidering Tonotopic Maps in the Auditory Cortex and Lemniscal Auditory Thalamus in Mice." Front. Neural Circuits 11:14 doi:10.3389/fncir.2017.00014
- ^ Bendor, D; Wang, X (2005). “The neuronal representation of pitch in primate auditory cortex.”. Nature 436 (7054): 1161-5. doi:10.1038/nature03867.
- ^ Zatorre, RJ (2005). “Neuroscience: finding the missing fundamental”. Nature 436 (7054): 1093-4. doi:10.1038/4361093a.
- ^ Access : : Nature
- ^ Klinke, Rainer; Kral, Andrej; Heid, Silvia ; Tillein, Jochen ; Hartmann , Rainer (Sep. 10, 1999). “Recruitment of the auditory cortex in congenitally deaf cats by long-term cochlear electrostimulation”. Science 285 (5434): 1729-33. doi:10.1126/science.285.5434.1729.
- ^ Strickland (Winter 2001). “Music and the brain in childhood development”. Childhood Education 78 (2): 100-4.
- ^ Bertrand, O.; Tallon-Baudry, C.; Fischer, C.; and Pernier, J.. Object representation and gamma oscillations.
- ^ Knief, A.; Schulte, M.; Fujiki, N.; and Pantev, C.. Oscillatory Gamma band and Slow brain Activity Evoked by Real and Imaginary Musical Stimuli.
- ^ Abbott, Alison Music, maestro, please! Nature v. 416 no. 6876 (March 7 2002)
- ^ Petr Janata et al. The Cortical Topography of Tonal Structures Underlying Western Music. Science, Vol 298, Issue 5601, 2167-2170 , 13 December 2002
- ^ Cassel, Topography of projections from the medial prefrontal cortex to the amygdala in the rat. Brain Res Bull. 1986 Sep;17(3):321-33