
線条体
| 脳: 線条体 、纹状体 | |
|---|---|
 ヒト脳内での線条体の位置。赤色で示す領域が線条体。 左は側面から見た図。右は正面から見た図。  | |
| 名称 | |
| 日本語 | 線条体 、纹状体 |
| 英語 | striatum |
| ラテン語 | corpus striatum |
| 略号 | Str、CPu |
| 関連構造 | |
| 上位構造 | 終脳、大脳、大脳基底核 |
| 構成要素 | 背側線条体(尾状核、被殻、ストリオソーム、マトリックス)、腹側線条体(側坐核、嗅結節) |
| 関連情報 | |
| Brede Database | 階層関係、座標情報 |
| NeuroNames | 関連情報一覧 |
| NIF | 総合検索 |
| MeSH | Corpus+Striatum |
| グレイ解剖学 | 書籍中の説明(英語) |
線条体(せんじょうたい、striatum)は、終脳の皮質下構造であり、大脳基底核の主要な構成要素のひとつである。線条体は運動機能への関与が最もよく知られているが、意思決定などその他の神経過程にも関わると考えられている。線条体は、新線条体(または背側線条体)と腹側線条体に区分されるが、単に「線条体」と言った場合には新線条体のことを指す場合が多い。線条体 striatum という名称は、ヒト新線条体の尾状核と被殻が内包(大脳新皮質や視床からの軸索線維の束であり白質)によって分断される場所で、互いに連絡している部分が線条 stria として見えることから命名された[1]。
新線条体
[編集]新線条体(しんせんじょうたい neostriatum)は、背側線条体とも呼ばれ、被殻(ひかく putamen)と尾状核(びじょうかく caudate nucleus)から構成されている(このため両者の総称として新線条体のことを Caudate-Putamen, CPu とも称する)。単に線条体と言った場合に、この新線条体のことを指す場合が多い。これとは別に新線条体(被殻と尾状核)は、ストリオソーム(striosome、またはパッチ patch とも呼ばれる)とマトリックス(matrix)の二領域に分けられ、マトリックスの中にモザイク状のストリオソームが埋め込まれるという特殊な細胞構築を示す。両者は、辺縁皮質と、大脳新皮質からそれぞれ興奮性入力を受けることが知られている。
神経細胞
[編集]線条体を構成するニューロンの大部分は、投射細胞であるGABA作動性の中型有棘細胞であるが、その他の抑制性神経細胞も存在する。新線条体を構成する神経細胞については、主にげっ歯類を用いた研究に基づき次のように分類されている。
投射神経細胞
[編集]新線条体の投射神経細胞は 中型有棘神経細胞(中型有棘ニューロン、medium-sized spiny neuron, または medium spiny neuron, MSN)と呼ばれる。GABA作動性で、樹状突起に密な棘突起をもち、細胞体の大きさが中程度(20μmほど)。ラットの新線条体の神経細胞の95%を占めると言われる。GABAの他に、エンケファリンやダイノルフィンなどのオピオイドや、P物質などのペプチドを産生する。生体内での発火頻度が低いのが特徴とされる。ウレタン麻酔下では膜電位がDOWN状態とUP状態のふたつの状態を1Hz前後で遷移することが知られる。DOWN状態では、内向き整流性カリウムチャネルKIR2の働きによって、過分極側でカリウムイオンの流入が生じる内向き整流性を顕著に示し、カリウムイオンの膜電位が反転電位付近の値をとる[2]。
いくつかの矛盾する報告もあるものの、現在では線条体の投射ニューロン(中型有棘ニューロン)は、直接路と間接路のふたつの経路を構成する2群に分類できるという見方が有力である。下記、出力の項を参照。
抑制性神経細胞
[編集]- アセチルコリン作動性抑制性細胞(cholinergic interneuron)
- アセチルコリン作動性。形態学的には大型無棘細胞(large aspiny neuron)と呼ばれ、電気生理学的にはTAN(tonically active neuron)と呼ばれているものがこれに相当すると考えられている。行動選択において重要な状況、もしくは報酬に関わる事象が発生した時に、この神経細胞は一時的に発火を停止する。
- GABA作動性抑制性神経細胞(GABAergic interneuron)
- パルブアルブミン陽性抑制性神経細胞(parvalbumin-positive interneuron)
- 電気生理学的には fast-spiking (FS) 細胞と呼ばれる。カルシウム結合蛋白質の一種であるパルブアルブミン陽性である。また線条体の細胞の中で、最もGABAの産生量が多い細胞と考えられている。
- ソマトスタチン作動性抑制性神経細胞(somatostatinergic interneuron)
- カルレチニン陽性抑制性神経細胞(calretinin-positive interneuron)
- 電気生理学的には low-thredhold spiking (LTS) 細胞 と呼ばれる。カルシウム結合蛋白質の一種であるカルレチニン陽性である。
- パルブアルブミン陽性抑制性神経細胞(parvalbumin-positive interneuron)
入力
[編集]- 皮質線条体入力(corticostriatal input)はグルタミン酸作動性であり、運動に関わる新皮質領野などからの入力が多いとされるが、辺縁皮質の他、ほとんどすべての大脳皮質領野からの入力が存在しており、大脳基底核が運動だけに関わっているわけではないという考えの根拠となっている。それぞれの皮質領野からの入力は線条体の内部で特定の領域に軸索を展開するため、線条体内部には領域によってゆるやかな機能分化があるとされる。大脳基底核における機能的平行ループの概念[3]もこれに基づいている。主に大脳皮質の出力層である5層の錐体細胞が、皮質線条体入力を供給している。
- 視床線条体入力(thalamostriatal input)もグルタミン酸作動性であるが、視床核の中でも非特殊核と呼ばれる核からの入力が中心を占める。さらにその中でもCM-PF複合体(centromedian parafascicular nuclear complex)からの入力が主である。
- 黒質線条体入力(nigrostriatal input)は黒質緻密部のA9細胞集団からのドーパミン作動性繊維である。
- そのほかに淡蒼球外節からのGABA作動性入力[4][5]、視床下核からの入力[6]も存在が報告されている。
出力
[編集]線条体の投射神経細胞は中型有棘神経細胞(medium-sized spiny neuron, または medium spiny neuron)である。いくつかの矛盾する報告もあるものの、現在では線条体の投射ニューロン(中型有棘ニューロン)は、直接路と間接路のふたつの経路を構成する、重複の小さい2群に分類できるという見方が有力である。ラットを用いた研究では中型有棘ニューロンは線条体の神経細胞の95%を占め、そのうち直接路と、間接路を構成するニューロンの比はほぼ同数であるという。
直接路を構成する中型有棘ニューロン
[編集]- 大脳基底核の出力核である、淡蒼球内節や黒質網様部へ投射。
- ドーパミンD1受容体(Drd1)陽性であり、中脳からのドーパミン入力によって、興奮性が高まる。
- P物質前駆体(Tac1)陽性、ダイノルフィン前駆体(Pdyn)陽性、アデノシンA1受容体陽性である。
- 従来は、途中の淡蒼球外節へは出力しないと考えられてきたが、近年の単一神経細胞の軸索追跡研究の結果、淡蒼球外節へも軸索側枝を出すものが多いという報告[7]もある。
間接路を構成する中型有棘ニューロン
[編集]- 淡蒼球外節へ投射する(淡蒼球内節や黒質へは投射しない)。
- ドーパミンD2受容体(Drd2)陽性であり、中脳からのドーパミン入力によって、興奮性が低下する。
- エンケファリン前駆体(Penk1)陽性、アデノシンA2A受容体陽性である。
発生
[編集]新線条体の中型有棘神経細胞は、外側基底核原基(lateral ganglionic eminence, LGE)に由来する。またアセチルコリン作動性抑制性神経細胞は、淡蒼球の原基である内側基底核原基(medial ganglionic eminence, MGE)からの細胞移動によって供給される。
ストリオソーム領域の中型有棘神経細胞は、マトリックスの中型有棘神経細胞よりも最終分裂日が早いことがげっ歯類を用いた研究[8]で明らかになっている。中脳由来のドーパミン含有線維(黒質線条体入力)は、発達期にはストリオソーム領域の中に高密度に分布して島状に見えることから、その構造を指してドーパミン・アイランドと呼ばれる。歴史的には発達期のみに見られるドーパミン・アイランドの概念が先にあって、後に成体脳におけるストリオソーム構造が見つかり、ドーパミン・アイランドと一致することが分かった[9][10]。
腹側線条体
[編集]腹側線条体(ふくそくせんじょうたい ventral striatum)は、側坐核(そくざかく accumbens nucleusまたはnucleus accumbens(ラテン語))、嗅結節(きゅうけっせつ olfactory tubercle、または tuberculum olfactorium(ラテン語))などを含む。種々の薬物中毒に関わっているとされる。
画像
[編集]-
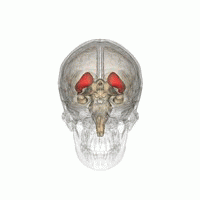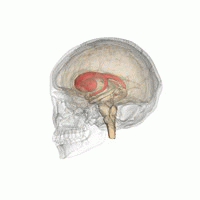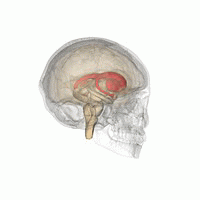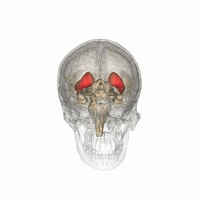 線条体の位置を様々な角度から眺めた動画。赤色で示す領域が線条体。(画像出典:Anatomography)
線条体の位置を様々な角度から眺めた動画。赤色で示す領域が線条体。(画像出典:Anatomography) -



脚注
[編集]- ^ 標準生理学第6版 医学書院
- ^ Nisenbaum ES, Wilson CJ (1995). “Potassium currents responsible for inward and outward rectification in rat neostriatal spiny projection neurons.”. J Neurosci 15 (6): 4449-63. PMID 7790919.
- ^ Alexander GE, Crutcher MD, & DeLong MR (1990). “Basal ganglia-thalamocortical circuits: parallel substrates for motor, oculomotor, "prefrontal" and "limbic" functions.”. Prog Brain Res 85: 119-146. PMID 2094891.
- ^ Beckstead RM (1983). “A pallidostriatal projection in the cat and monkey.”. Brain Res Bull 11 (6): 629-632.. PMID 6661668.
- ^ Staines WA, Atmadja S, & Fibiger HC (1981). “Demonstration of a pallidostriatal pathway by retrograde transport of HRP-labeled lectin.”. Brain Res 206 (2): 446-450.. PMID 7214143.
- ^ Kita H & Kitai ST (1987). “Efferent projections of the subthalamic nucleus in the rat: light and electron microscopic analysis with the PHA-L method.”. J Comp Neurol 260 (3): 435-452.. PMID 2439552.
- ^ Lévesque M, Parent A. (2005). “The striatofugal fiber system in primates: a reevaluation of its organization based on single-axon tracing studies.”. Proc Natl Acad Sci U S A 102 (33): 11888-93. PMID 16087877.
- ^ van der Kooy D, Fishell G (1986). “Neuronal birthdate underlies the development of striatal compartments.”. Brain Res 401 (1): 155-61. doi:10.1016/0006-8993(87)91176-0. PMID 3028569.
- ^ Graybiel AM, Pickel VM, Joh TH, Reis DJ, Ragsdale CW Jr. (1981). “Direct demonstration of a correspondence between the dopamine islands and acetylcholinesterase patches in the developing striatum.”. Proc Natl Acad Sci U S A. 78 (9): 5871-5.. PMID 6117860.
- ^ Herkenham M, Pert CB. (1981). “Mosaic distribution of opiate receptors, parafascicular projections and acetylcholinesterase in rat striatum.”. Nature 291 (5814): 415-8.. PMID 6165892.
関連項目
[編集]外部リンク
[編集]
| 国立図書館 | |
|---|---|
| その他 | |
この項目は、医学に関連した書きかけの項目です。この項目を加筆・訂正などしてくださる協力者を求めています(プロジェクト:医学/Portal:医学と医療)。 |