
ラビリンチュラ綱
| ラビリンチュラ綱 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 アプラノキトリウム属 (ラビリンチュラ目)
| |||||||||||||||
| 分類 | |||||||||||||||
| |||||||||||||||
| 学名 | |||||||||||||||
| Labyrinthulea Cavalier-Smith, 1986 (Labyrinthulomycetes Arx, 1970) | |||||||||||||||
| 和名 | |||||||||||||||
| ラビリンツラ綱、ラビリンチュラ菌綱、ラビリンツラ菌綱、 ラビリンチュラ類 (広義) | |||||||||||||||
| 英名 | |||||||||||||||
| labyrinthulomycetes, labyrinthulids (s. l.) | |||||||||||||||
| 下位分類 | |||||||||||||||
ラビリンチュラ綱、ラビリンツラ綱 (学名: Labyrinthulea[注 1])、ラビリンチュラ菌綱、ラビリンツラ菌綱 (学名: Labyrinthulomycetes[注 2]) は、ストラメノパイル (不等毛類) に属する菌類様の原生生物の1群である。栄養体は基本的に単細胞性であるが、ラビリンチュラ属 (ラビリンツラ属[1]、ラビリンスラ属[2]) は特異な網状の群体を形成する。ふつう細胞は薄い鱗片が重なった外被で覆われ、また外質ネットとよばれる仮根または糸状仮足様の構造を伸ばしている (右図)。ラビリンチュラ綱の中で、細胞の1または数箇所から仮根状の外質ネットを伸ばしている種は、しばしばヤブレツボカビ類 (英: thraustochytrids) とよばれる[3][4]。海産種が多く知られるが、淡水や陸上に生育する種もいる。基本的に吸収栄養性であり、植物遺骸などを細胞外消化・吸収する腐生性、または生きた生物から栄養を吸収する共生性 (寄生性を含む) である。多価不飽和脂肪酸 (DHAなど) や炭化水素 (スクアレンなど) を多く産生する種が知られており、応用研究が盛んで一部は実用化されている。
一般名としてはラビリンチュラ類、ラビリンツラ類 (英: labyrinthulids) とよばれるが[3][4]、この名はラビリンチュラ属のみを意味することもある[5] (以下ではラビリンチュラ綱全体に対する一般名として「ラビリンチュラ類」を用いている)。またラビリンチュラ類は光合成能を欠き、藻類に分類されたことはないが、不等毛藻 (オクロ植物門;珪藻や褐藻を含む) に比較的近縁であるため、藻類とよばれることがある[5]。
特徴
[編集]形態
[編集]栄養体 (通常時の体) は基本的に単細胞性であるが (下図1)、ラビリンチュラ属 (Labyrinthula) は特異な網状の変形体様群体を形成する[1][5][6][7] (下図2; 下記参照)。ソロディプロフリス属 (Sorodiplophrys) の栄養体は単細胞性であるが、胞子形成時に細胞が集合して細胞性粘菌の累積子実体によく似た子実体を形成する[8][9]。


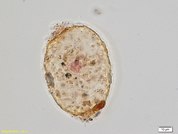
細胞はふつう薄い (厚さ 2–3 nm) 盤状の鱗片が重なり合った外被構造で囲まれている[1][4][5][7][10] (上図1b)。この鱗片は硫酸化多糖 (ガラクトース、キシロース、フコースなどに富み、系統群によって組成が異なる[10][11]) からなり、ゴルジ体で形成される[5][7][12]。一部の種は、生活環の一時期に鱗片を欠くアメーバ状細胞 (amoeboid cell) を形成する[5]。例外的に、アンフィトレマ科の種は両端に開口部をもつ厚くゆるい殻 (ロリカ) を形成する[13][14] (上図1c)。またラビリンチュラ類の中には、細胞外多糖 (exopolysaccharide) を多く分泌するものもいる[5]。細胞外多糖はグルコースやガラクトースを多く含む[15]。
細胞の一端または複数箇所から、外質ネット[3][4][16] (外質網[17]、網状構造[1]; ectoplasmic net, – network, – thread, – filament) とよばれる構造が生じている[5][7][18]。外質ネットはしばしば分枝する仮根または糸状仮足様の構造であり、細胞外被を欠き、生体膜に包まれているがリボソームや細胞小器官をほとんど含まない[3][6][7][19]。ただし、内部にしばしば小胞が存在し、またアクチン繊維を含む[10]。外質ネットはふつう細胞表層に存在するボスロソーム[4][16] (bothrosome; 外質網形成体[17]、サゲノジェネトソーム sagenogenetosome, sagenetosome[6], sagenogen[20]) とよばれる細胞小器官から生じている[5][18][19][21]。ボスロソームは小胞体が密集し電子密度が高い物質が付随している。ただしアンフィフィラ属 (Amphifila) などでは外質ネットは存在するが、ボスロソームは確認されていない[22]。外質ネットは基質に付着し、酵素を分泌 (細胞外消化する)、栄養物を吸収すると考えられている[5][7][12][19]。


ラビリンチュラ属では、細胞から生じた外質ネットが細胞全体を包み、さらに多数の細胞の外質ネットが融合して網状の変形体様の構造を形成する[3][4][5][7][10][12] (上図2)。そのためラビリンチュラ属は net slime molds や slime nets ともよばれ[10][20]、またこのような構造は網状変形体 (net plasmodium, filoplasmodium) ともよばれていた[6]。ラビリンチュラ属の属名の語源である labyrinthus はラテン語で「迷路」を意味し、このような網状構造の形を示している[17]。この網状構造では多数の細胞が共通の生体膜で包まれているが、各細胞はボスロソームによって区切られている。各細胞は、この共通基質中をアクチン・ミオシンが関与する滑走運動をする (100 µm/分に達する)[3][20]。アプラノキトリウム属は基本的に単細胞性であるが、ラビリンチュラ属に似た外質ネットに沿った滑走運動能をもつ[7]。またヤブレツボカビ科の中には、生活環の一時期に運動能をもつアメーバ細胞を形成するものもいる[10]。

細胞は基本的に単核性 (核を1個のみもつ) であり、ふつう核に接して中心体が存在する[5]。ミトコンドリアのクリステは管状[5][10]。ゴルジ体が存在し、また核に近接して滑面小胞体に囲まれたリボソームを欠く領域 (paranuclear body) をもつことが多い[5]。細胞質中にはしばしば多量の脂質顆粒が存在し、大きな脂質顆粒が細胞の大部分を占めていることもある[5][10] (右図3)。脂質顆粒は透明のものから、カロテノイドを含んで橙色から赤色を呈するものまである。乾燥重量あたりの総脂肪酸量は8%から82%に達する[5]。
ラビリンチュラ類は、色素体に関連する細胞小器官をもたない。例外的に、アンフィトレマ科の種は緑藻のクロレラ (トレボウクシア藻綱) を細胞内共生させている[23]。
生殖
[編集]
二分裂、遊走子 (鞭毛をもつ胞子)、不動胞子、アメーバ細胞形成などによる無性生殖が報告されている[3][4][5][10][16] (右図4)。基本的に分実単心性であり、外質ネット以外の細胞全体が胞子嚢になり、細胞外被で囲まれて胞子を形成、細胞外被が崩壊することで胞子が放出される[1][5][10]。また、胞子放出後に胞子嚢内に残った部分 (原基体 proliferous body) が成長し、再び栄養細胞になるものも知られている[16][24]。ラビリンチュラ属では、シスト形成が知られている[6]。

遊走子は細胞側面 (腹面) から前後に伸びる2本の不等鞭毛をもち、前鞭毛には管状小毛が付随している[5][6] (上図5)。ラビリンチュラ属の遊走子は眼点をもち、走光性を示すことが報告されている[1][25]。遊走子は、栄養細胞と同様に鱗片で覆われていることがある[10]。多くのストラメノパイルの鞭毛の基部に存在するらせん構造は存在せず、かわりに円錐状の構造が報告されている[10][26]。
ソロディプロフリス属は陸上生であり、胞子形成時に細胞が集合して細胞性粘菌でみられる子実体によく似た子実体 (累積子実体) を形成する[8][9]。
有性生殖については明らかではない[10]。ラビリンチュラ属の遊走子形成時の第1分裂においてシナプトネマ構造が観察されており (染色体数は n = 9)、これが減数分裂であることが示唆されている[6][27]。ただし、細胞質融合や核融合の時期は明らかではない。一方で、ヤブレツボカビ類において鞭毛細胞の合体が報告されているが、その後の経過については不明である[28] (右図4V)。また一部の種で休眠胞子形成が知られているが、これが配偶子合体の結果であるか否かは定かではない[7][19]。
生態
[編集]ラビリンチュラ類の既知種の多くは海から汽水域にかけて生育し、また高塩分環境からも報告されている[5][10]。熱帯から極地まで分布し、沿岸域に多いが、外洋の深海域からも報告されている[5][7][10]。その生物量も多く、瀬戸内海ではラビリンチュラ類の生物量が細菌の生物量の3.5–41%に相当することが報告されている[3]。水底堆積物、マングローブ林、河口など一般的に有機物が豊富な環境に多い。浮遊有機物に付着したプランクトン性のものもいる[10]。また淡水や陸上に分布しているものもおり、例えばアンフィトレマ科の種は高層湿原に[13][29]、ディプロフリス属は一般的な湖沼に[29][30]、ソロディプロフリス属は土壌に[8][注 3]生育している。環境DNAに基づく研究からは、淡水や陸上域には未知のラビリンチュラ類が多く存在することが示唆されている[7]。またこのような研究からは、これまでラビリンチュラ類の報告がない嫌気的な堆積物などの環境にも、ラビリンチュラ類が生育していることが示唆されている[10]。
基本的に吸収栄養生物 (細胞膜を通して可溶性有機物を吸収して利用する生物) であり、動植物遺骸など生きていない有機物を細胞外消化、吸収する腐生性のものが多く知られている[1][3][5][10][20]。このようなラビリンチュラ類は、特に海洋生態系において分解者として重要な生物群であると考えられている[10][31][32]。特に細菌などでは分解が難しい陸上植物由来の有機物 (セルロースなど) の分解において、ラビリンチュラ類が重要な役割を担っていることが示唆されている[3][32]。さらにラビリンチュラ類は細菌にくらべて大型であるため、腐植物から高次捕食者へ直接エネルギーや物質が流れる経路にもなると考えられている (細菌-細菌捕食者の段階を省略する)[3][32]。またラビリンチュラ類は、死骸だけではなく生きている微生物を分解・吸収することもある。培養下では、細菌や酵母、糸状菌、珪藻、糸状藻、さらに他のラビリンチュラ類を分解することが報告されている[10]。
ラビリンチュラ類は、株によってアガラーゼ、カラギナーゼ、アルギン酸リアーゼ、ペクチナーゼ、キシラナーゼ、キチナーゼ、セルラーゼ、アミラーゼ、α-グルコシダーゼ、プロテアーゼ、ゼラチナーゼ、ウレアーゼ、リパーゼ、ホスファターゼなどの分解酵素活性をもつ[5]。また、遊走子がペクチンに対する走化性を示すことが報告されている[3]。少なくとも一部の種は、極めて強固な物質である花粉のスポロポレニン分解能をもつ[6]。ヤブレツボカビ類を野外から単離する際には、一般的に花粉を餌とする方法が用いられている[20]。
さらにラビリンチュラ類の中には、生きている生物に共生して栄養を得る共生性 (寄生性を含む) のものも多いと考えられている[1][7]。さまざまな海藻や海草の表面や内部からラビリンチュラ類が見つかっており、これらは共生性 (相利共生、片利共生、または寄生) である可能性がある[10]。ラビリンチュラ属の中には、海草のアマモに寄生する種 (Labyrinthula zosterae) や、陸生植物である芝に寄生する種 (Labyrinthula terrestris) も知られている[33][34] (下記参照)。また珪藻の細胞内に外質ネットまたは本体を侵入させる寄生性の種も報告されており (Phycophthorum)、このようなラビリンチュラ類が海洋に広く生育していることが示唆されている[35]。ラビリンチュラ属の1種が、アーバスキュラー菌根を形成する菌類であるグロムス類の胞子に寄生している例も報告されている[36]。他にもアメーバ類に細胞内共生する例や[37]、海綿、サンゴ、ウニ、ホヤ、ニジマス、扁形動物、さまざまな軟体動物 (タコやイカ、ウミウシ類、アワビ、二枚貝) に共生 (寄生) している例が知られている[5][10][38][39]。ただし大型の生物に対する共生性とされる例の中には、実際にはそこに付着している微生物を分解・吸収していることもある[10]。
少なくとも一部の種では、細菌などを取り込む食作用能をもつことが示唆されている[13][40]。
ラビリンチュラ類は光合成能をもたないが、アンフィトレマ科の生物は緑藻のクロレラ (トレボウクシア藻綱) を細胞内共生させており、おそらく共生藻の光合成による有機物を得ていると考えられている[23][41]。
ラビリンチュラ類に寄生するウイルス (1本鎖RNAウイルス、2本鎖DNAウイルス) が見つかっている[3][18]。上記のようにラビリンチュラ類は海洋生態系において分解者として重要な要素であると考えられているが、その増減にウイルスが大きく影響している可能性がある。また共生性のラビリンチュラ類の場合、ラビリンチュラ類が宿主に対するウイルスの媒介者になっている可能性もある[10]。
人間との関わり
[編集]1930年代に北大西洋両岸でアマモ群落 (アマモ場) の大規模な減少が起こり、魚介類など水産資源に大きな影響を与えた。このアマモ衰弱病 (eelgrass wasting disease) の原因はラビリンチュラ属の1種 (Labyrinthula zosterae) の寄生によるものとされるが、その詳しい関係は明らかではない[3][6][10][20][42][43]。またラビリンチュラ属の別の1種 (Labyrinthula terrestris) は芝 (ホソムギ、ハイコヌカグサ、イヌコヌカグサ、オオスズメノカタビラ) に寄生し、ゴルフ場の芝をパッチ状に枯らすことがある (ラピッドブライト病 rapid blight disease)[3][34][44]。ホンビノスガイに寄生する Mucochytrium quahogii は QPX (Quahog Parasite Unknown) とよばれ、北米で大きな被害を与えている[45][46]。また Aplanochytrium haliotidis はアワビ類に寄生して被害を与える[47]。
ラビリンチュラ類は脂質を多く生成するため、有用物質生産の観点から注目されている。ラビリンチュラ類はドコサヘキサエン酸 (DHA) やドコサペンタエン酸 (DPA)、エイコサペンタエン酸 (EPA)、アラキドン酸 (ARA) など種によって異なる高度不飽和脂肪酸を多く生成する[5]。動物由来ではないという付加価値をもつため、ラビリンチュラ類由来の高度不飽和脂肪酸が複数の企業から販売されている[5][48][49][50]。ラビリンチュラ類は種によってアスタキサンチンやカンタキサンチン、β-カロテンなどのカロテノイドを産生し、その応用も試みられている[5][48][51]。またラビリンチュラ類の中には、炭化水素であるスクアレンを多く産生する株も知られている (オーランチオキトリウム属の1種)[52]。スクアレンは健康食品や医薬品に利用されており、またバイオ燃料としての可能性もあるため、その観点でのラビリンチュラの研究も進められている[5][48]。
系統と分類
[編集]ラビリンチュラ属は網状の変形体様構造を形成し、細胞はその中で滑走運動をする (上記参照)。このような特徴から、ラビリンチュラ属は粘菌 (広義の変形菌)[注 4] や肉質虫 (広義のアメーバ類) に分類されていた[1][6][16][53][54][55]。さらにその特異性から、原生動物や原生生物の独立の門 (ラビリンチュラ門) に分類されることもあった[17][53][56]。一方、ヤブレツボカビ類はツボカビ類に似た体 (仮根様構造が生じた球状の細胞) をもつものの、卵菌に似た鞭毛細胞を形成することから、卵菌綱に分類されていた[5][10][57]。
やがて1970年代以降の電子顕微鏡を用いた微細構造学的研究から、ラビリンチュラ属とヤブレツボカビ類は様々な特徴 (鱗片が重なった外被、ボスロソームから生じる外質ネットなど) を共有しており、またストラメノパイル (不等毛類) の特徴 (管状小毛を伴う前鞭毛など) をもつことが明らかとなった。そのため、ラビリンチュラ属とヤブレツボカビ類がストラメノパイル内で1つの系統群を構成していると考えられるようになった[10][12][58][59]。その後20世紀末以降の分子系統学的研究からも、この考えは支持されている[11][60]。また微細構造や分子系統学的研究から、糸状仮足をもつアメーバ類 (糸状仮足類) として扱われていたいくつかの原生生物 (ディプロフリス属、アンフィトレマ科など) も、ラビリンチュラ類に属することが示されている[13][61][62]。2020年現在では、これらの生物を合わせてラビリンチュラ綱[63] (ラビリンツラ綱[64] Labyrinthulea[注 1];ラビリンチュラ菌綱[16]、ラビリンツラ菌綱[65] Labyrinthulomycetes[注 2]) に分類することが一般的となっている。さらに独立の門 (ラビリンチュラ門[63]、ラビリンツラ門[64] Labyrinthomorpha[10][注 1];ラビリンチュラ菌門[16]、ラビリンツラ菌門[65] Labyrinthulomycota[10][66][注 2]) として扱われることもある。一方で多数の遺伝子塩基配列に基づく系統解析からは、捕食性鞭毛虫である Pseudophyllomitus がラビリンチュラ類に近縁であることが示唆されており、両者をあわせてサゲニスタ門または亜門[67] (Sagenista) に分類することもある[68][注 5]。また環境DNAの研究からは、実体不明であるいくつかの系統群 (MAST-4, 7, 8, 9, 10, 11, 20) がラビリンチュラ類に近縁であることが示唆されている[70]。ストラメノパイルの中には、菌類的な吸収栄養生物であるグループとして他に卵菌類が存在するが、ラビリンチュラ類と卵菌類は近縁ではない[10][11][60]。
ストラメノパイルの中では、不等毛藻は紅藻由来の色素体をもつが、ラビリンチュラ類や卵菌、オパリナ、ビコソエカ類などそれ以外のグループは全て従属栄養性であり色素体をもたない。不等毛藻がもつ色素体の起源は、クリプト藻やハプト藻がもつ色素体の起源と同一であるとする仮説があり (クロミスタ仮説)、この仮説ではラビリンチュラ類の祖先も紅藻由来の色素体をもっていたがそれを二次的に失ったとされる[67]。しかし2020年現在、ラビリンチュラ類が色素体をもっていたことを明らかに支持する証拠は得られておらず、ラビリンチュラ類が光合成生物に由来したか否かは明らかではない[10]。
ラビリンチュラ綱の中には、ラビリンチュラ目とヤブレツボカビ目が認識されている[1][3][11]。ヤブレツボカビ類のいくつかの属 (Schizochytrium, Ulkenia など) について分類学的再編成が行われ[16][71][72][73]、その過程でヤブレツボカビ類は単系統群ではないことが示唆されており、一部の属 (Aplanochytrium, Oblongichytrium) はヤブレツボカビ目から除かれるようになった[10]。また糸状仮足をもつアメーバ類として扱われていたいくつかの生物が、ラビリンチュラ綱を構成する別の系統群 (アンフィトレマ目、アンフィフィラ目) であることが示唆されている[13][62]。このようなラビリンチュラ綱内の分類については、分子系統学的解析や生活環のパターン、微細構造、多価不飽和脂肪酸やカロテノイドの組成などに基づいて行われている[3][5][71][72][74]。また環境DNAの研究からは、ラビリンチュラ類の中に実体不明の系統群が多数存在することが示されている[10][75]。既知種の系統仮説の一例と、2021年現在の分類体系の一例を下に示す (下図、下表)。
| |||||||||||||||||||||||||||||||||||||||
| ラビリンチュラ綱内の系統仮説の一例[10][13][62][74][76][77] |
ラビリンチュラ綱の属までの分類体系の一例 (2021年現在)[10][66][76]:綱から科の学名は国際動物命名規約上のものを主とし、[ ] 内に国際藻類・菌類・植物命名規約上のものを示す。
|
脚注
[編集]注釈
[編集]- ^ a b c 国際動物命名規約における学名である。
- ^ a b c 国際藻類・菌類・植物命名規約における学名である。
- ^ ソロディプロフリス属はウシやウマの糞から単離されている[9]。
- ^ 広義の変形菌門の中の水生変形菌綱 (Hydromyxomycetes) やラビリンチュラ菌綱 (Labyrinthulomycetes) に分類されていた[6][53]。
- ^ サゲニスタ門が提唱された当初はビコソエカ類など他の生物群も含んでいた[69]。
- ^ Labyrinthulea Levine & Corliss, 1963 としている例もある[56]。
- ^ "アプラノキトリウム目" ("Aplanochytriales"; 非正式名) として独立目としている例もある[44]。
- ^ 外質ネットを全く欠くことから他と分けられている。DNAデータが得られておらず、その系統的位置は明らかではない[10]。ヤブレツボカビ科に含めることもある[16]。
- ^ ソロディプロフリス科に分類している例もある[44]。
- ^ ステラルキトリウム科をラビリンチュラ目に分類している例もある[10][70]。
- ^ 環境DNAのみが知られていた LAB 1/6/8 とよばれる系統群に相当する[75][77]。
出典
[編集]- ^ a b c d e f g h i j 徳増 征二 (2005). “ラビリンツラ菌門”. In 杉山 純多 (編). バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統. 裳華房. pp. 188–189. ISBN 978-4785358273
- ^ “ラビリンスラ”. ブリタニカ国際大百科事典 コトバンク. 2020年11月11日閲覧。
- ^ a b c d e f g h i j k l m n o p q 本多 大輔 (2012). “ラビリンチュラ類”. In 渡邉 信 (監). 藻類ハンドブック. 株式会社エヌ・ティー・エス. pp. 73-76. ISBN 978-4864690027
- ^ a b c d e f g 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “ラビリンツラ類”. 岩波 生物学辞典 第5版. 岩波書店. p. 1437. ISBN 978-4000803144
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad Marchan, L. F., Chang, K. J. L., Nichols, P. D., Mitchell, W. J., Polglase, J. L. & Gutierrez, T. (2018). “Taxonomy, ecology and biotechnological applications of thraustochytrids: A review”. Biotechnology Advances 36: 26-46. doi:10.1016/j.biotechadv.2017.09.003.
- ^ a b c d e f g h i j k ジョン・ウェブスター (著) 椿 啓介・三浦宏一郎・山本昌木 (訳) (1985). “水生変形菌綱”. ウェブスター菌類概論. 講談社サイエンティフィク. pp. 20–23. ISBN 978-4061396098
- ^ a b c d e f g h i j k l Beakes, G. W., Honda, D. & Thines, M. (2014). “Systematics of the Straminipila: Labyrinthulomycota, Hyphochytriomycota, and Oomycota”. In McLaughlin, D. J. & Spatafora, J. W.. THE MYCOTA, volume 7A. Systematics and Evolution Part A. Springer. pp. 39-97. doi:10.1007/978-3-642-55318-9_3
- ^ a b c Dykstra, M. J. & Olive, L. S. (1975). “Sorodiplophrys: an unusual sorocarp-producing protist”. Mycologia 67: 873-879. doi:10.2307/3758346.
- ^ a b c Tice, A. K., Silberman, J. D., Walthall, A. C., Le, K. N., Spiegel, F. W. & Brown, M. W. (2016). “Sorodiplophrys stercorea: another novel lineage of sorocarpic multicellularity”. Journal of Eukaryotic Microbiology 63: 623-628. doi:10.1111/jeu.12311.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am Bennett, R. M., Honda, D., Beakes, G. W. & Thines, M. (2017). “Labyrinthulomycota”. In Archibald, J. M., Simpson, A. G. B. & Slamovits, C. H.. Handbook of the Protists. Springer. pp. 507-542. ISBN 978-3319281476
- ^ a b c d Honda, D., Yokochi, T., Nakahara, T., Raghukumar, S., Nakagiri, A., Schaumann, K. & Higashihara, T. (1999). “Molecular phylogeny of labyrinthulids and thraustochytrids based on the sequencing of 18S RNA gene”. Journal of Eukaryotic Microbiology 46: 637-646. doi:10.1111/j.1550-7408.1999.tb05141.x.
- ^ a b c d Moss, S. (1986). “Biology and phylogeny of the Labyrinthulales and Thraustochytriales”. The Biology of Marine Fungi. Cambridge University Press. pp. 105-129. ISBN 978-0521308991
- ^ a b c d e f Gomaa, F., Mitchell, E. A. & Lara, E. (2013). “Amphitremida (Poche, 1913) is a new major, ubiquitous labyrinthulomycete clade”. PLoS One 8: e53046. doi:10.1371/journal.pone.0053046.
- ^ Lee, J. J., Leedale, G. F. & Bradbury, P., ed (2000). An Illustrated Guide to the Protozoa, Organisms Traditionally Referred to as Protozoa, or Newly Discovered Groups. Blackwell Publishers. p. 1064. ISBN 978-1891276224
- ^ Jain, R., Raghukumar, S., Tharanathan, R. & Bhosle, N.B. (2005). “Extracellular polysaccharide production by thraustochytrid protists”. Mar. Biotechnol. 7: 184–192. doi:10.1007/s10126-004-4025-x.
- ^ a b c d e f g h i 本多大輔 (2013). “1.8.5 ラビリンチュラ菌門”. In 日本菌学会 (編). 菌類の事典. 朝倉書店. pp. 39–41. ISBN 978-4254171471
- ^ a b c d 白山 義久 (編) (2000). “ラビリンツラ門”. バイオディバーシティ・シリーズ (5) 無脊椎動物の多様性と系統. 裳華房. pp. 80–81. ISBN 978-4785358273
- ^ a b c 高尾 祥丈, 長崎 慶三 & 本多 大輔 (2007). “ウイルス研究から見た海産原生生物ラビリンチュラ類の生態学的多様性”. 極限環境微生物学会誌 6: 68-73. doi:10.3118/jjse.6.68.
- ^ a b c d Porter, D. (1990). “Phylum Labyrinthulomycota”. In Margulis, L., Corliss, J. O., Melkonian, M. & Chapman, D. J.. Handbook of Protoctista. Jones and Bartlett. pp. 388–398. ISBN 978-0867200522
- ^ a b c d e f Alexopoulos, C. J., Mims, C. W. & Blackwell, M. (1996). “Phylum Labyrinthulomycota”. Introductory Mycology. John Wiley & Sons. Inc., New York. pp. 743–750. ISBN 978-0-471-52229-4
- ^ Moss, S. T. (1985). “An ultrastructural study of taxonomically significant characters of the Thraustochytriales and Labyrinthulales”. Bot. J. Linn. Soc. 91: 329–357. doi:10.1111/j.1095-8339.1985.tb01154.x.
- ^ Dykstra, M. J. & Porter, D. (1984). “Diplophrys marina, a new scale-forming marine protist with labyrinthulid affinities”. Mycologia 76 (4): 626-632. doi:10.1080/00275514.1984.12023892.
- ^ a b Lara, E. & Gomaa, F. (2017). “Symbiosis between testate amoebae and photosynthetic organisms”. Algal and Cyanobacteria Symbioses. pp. 399-419. doi:10.1142/9781786340580_0013
- ^ 本多 大輔 (2009年). “Laby Base”. 甲南大学. 2020年11月5日閲覧。
- ^ Perkins, F. O. & Amon, J. P. (1969). “Zoosporulation in Labyrinthula sp.; an electron microscope study”. The Journal of Protozoology 16: 235-257. doi:10.1111/j.1550-7408.1969.tb02265.x.
- ^ Barr, D. J. & Allan, P. M. (1985). “A comparison of the flagellar apparatus in Phytophthora, Saprolegnia, Thraustochytrium, and Rhizidiomyces”. Canadian Journal of Botany 63 (1): 138-154. doi:10.1139/b85-017.
- ^ Moens, P. B. & Perkins, F. O. (1969). “Chromosome number of a small protist: accurate determination”. Science 166 (3910): 1289-1291. doi:10.1126/science.166.3910.1289.
- ^ Ganuza, E., Yang, S., Amezquita, M., Giraldo-Silva, A. & Andersen, R. A. (2019). “Genomics, biology and phylogeny Aurantiochytrium acetophilum sp. nov.(Thraustrochytriaceae), including first evidence of sexual reproduction”. Protist 170: 209-232. doi:10.1016/j.protis.2019.02.004.
- ^ a b 月井 雄二 (2010). 淡水微生物図鑑 原生生物ビジュアルガイドブック. 誠文堂新光社. p. 50. ISBN 978-4416210048
- ^ 中山 剛 & 山口 晴代 (2018). プランクトンハンドブック 淡水編. 文一総合出版. p. 96. ISBN 978-4829981542
- ^ Bongiorni, L. (2012). “Thraustochytrids, a neglected component of organic matter decomposition and food webs in marine sediments”. In Raghukuma, C.. Biology of Marine Fungi. Prog. Mol. Subcell. Biol. 53.. Springer. pp. 1–13. ISBN 978-3-642-23342-5
- ^ a b c 中村 慶子 & 長沼 毅 (2007). “海洋環境におけるラビリンチュラ類の生態学的役割”. 日本海水学会誌 61: 316-320. doi:10.11457/swsj1965.61.316.
- ^ Bigelow, D. M., Olsen, M. W. & Gilbertson, R. L. (2005). “Labyrinthula terrestris sp. nov., a new pathogen of turf grass”. Mycologia 97: 185-190. doi:10.1080/15572536.2006.11832852.
- ^ a b Douhan, G. W., Olsen, M. W., Herrell, A., Winder, C., Wong, F. & Entwistle, K. (2009). “Genetic diversity in Labyrinthula terrestris, a newly emergent plant pathogen, and the discovery of new Labyrinthulid organism”. Mycol. Res. 113: 1192–1199. doi:10.1016/j.mycres.2009.08.002.
- ^ Hassett, B. T. (2020). “A widely distributed thraustochytrid parasite of diatoms isolated from the Arctic represents a gen. and sp. nov.”. The Journal of Eukaryotic Microbiology. doi:10.1111/jeu.12796.
- ^ Koske, R. E. (1981). “Labyrinthula inside the spores of a vesicular—arbuscular mycorrhizal fungus”. Mycologia 73 (6): 1175-1180. doi:10.1080/00275514.1981.12021451.
- ^ Dyková, I., Fiala, I., Dvořáková, H. & Pecková, H. (2008). “Living together: The marine amoeba Thecamoeba hilla Schaeffer, 1926 and its endosymbiont Labyrinthula sp”. European Journal of Protistology 44: 308-316. doi:10.1016/j.ejop.2008.04.001.
- ^ Azevedo, C. & Corral, L. (1997). “Some ultrastructural observations of a thraustochytrid (Protoctista, Labyrinthulomycota) from the clam Ruditapes decussatus (Mollusca, Bivalvia)”. Diseases of Aquatic Organisms 31: 73-78. doi:10.3354/dao031073.
- ^ Jones, G. M. & O’Dor, R. K. (1983). “Ultrastructural observations on a thraustochytrid fungus parasitic in the gills of squid (Illex illecebrosus Lesueur)”. J. Parasitol. 69: 903–911. doi:10.2307/3281055.
- ^ Raghukumar, S. (2002). “Bacterivory: a novel dual role for thraustochytrids in the sea”. Marine Biology 113: 165-169. doi:10.1007/BF00367650.
- ^ Gomaa, F., Kosakyan, A., Heger, T. J., Corsaro, D., Mitchell, E. A. & Lara, E. (2014). “One alga to rule them all: unrelated mixotrophic testate amoebae (Amoebozoa, Rhizaria and Stramenopiles) share the same symbiont (Trebouxiophyceae)”. Protist 165 (2): 161-176. doi:10.1016/j.protis.2014.01.002.
- ^ Muehlstein, L., Porter, D. & Short, F. (1991). “Labyrinthula zosterae sp. nov., the causative agent of wasting disease of eelgrass, Zostera marina”. Mycologia 83: 180–191. doi:10.1080/00275514.1991.12025994.
- ^ Brakel, J., Werner, F.J., Tams, V., Reusch, T.B.H. & Bockelmann, A.C. (2014). “Current European Labyrinthula zosterae are not virulent and modulate seagrass (Zostera marina) defense gene expression”. PLoS One 9. doi:10.1371/journal.pone.0092448.
- ^ a b c Preisfeld, A. (2012). “Labyrinthulomycota”. In Frey, W. (eds.). Syllabus of Plant Families. A. Engler's Syllabus der Pflanzenfamilien Part 1/1. Borntraeger. pp. 94-97. ISBN 978-3-443-01061-4
- ^ Bassim, S. & Allam, B. (2018). “SNP hot-spots in the clam parasite QPX”. BMC Genomics 19: 486. doi:10.1186/s12864-018-4866-8.
- ^ Geraci-Yee, S., Brianik, C. J., Rubin, E., Collier, J. L. & Allam, B. (2021). “Erection of a new genus and species for the pathogen of hard clams ‘Quahog Parasite Unknown’(QPX): Mucochytrium quahogii gen. nov., sp. nov.”. Protist 172 (1): 125793. doi:10.1016/j.protis.2021.125793.
- ^ Bower, S. M., McLean, N. & Whitaker, D. J. (1989). “Mechanism of infection by Labyrinthuloides haliotidis (Protozoa, Labyrinthomorpha), a parasite of abalone (Haliotis kamtschatka) (Mollusca, Gastropoda)”. Journal of Invertebrate Pathology 53: 401–409. doi:10.1016/0022-2011(89)90106-7.
- ^ a b c Aasen, I. M., Ertesvåg, H., Heggeset, T. M. B., Liu, B., Brautaset, T., Vadstein, O. & Ellingsen, T. E. (2016). “Thraustochytrids as production organisms for docosahexaenoic acid (DHA), squalene, and carotenoids”. Applied Microbiology and Biotechnology 100: 4309-4321. doi:10.1007/s00253-016-7498-4.
- ^ Winwood, R. J. (2013). “Recent developments in the commercial production of DHA and EPA rich oils from micro-algae”. OCL 20 (6): D604. doi:10.1051/ocl/2013030.
- ^ “なぜlife's製品なのか?”. life'sDHA. 2020年8月15日閲覧。
- ^ Aki, T., Hachida, K., Yoshinaga, M., Katai, Y., Yamasaki, T., Kawamoto, S., ... & Ono, K. (2003). “Thraustochytrid as a potential source of carotenoids”. Journal of the American Oil Chemists' Society 80: 789-794.
- ^ Kaya, K., Nakazawa, A., Matsuura, H., Honda, D., Inouye, I. & Watanabe, M.M. (2011). “Thraustochytrid Aurantiochytrium sp. 18W-13a accummulates high amounts of squalene”. Biosci. Biotechnol. Biochem. 75: 2246–2248. doi:10.1271/bbb.110430.
- ^ a b c 杉山純多 (2005). “菌類の多様性と分類体系”. In 杉山純多 (編). バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統. 裳華房. pp. 38–39. ISBN 978-4785358273
- ^ 井上 浩, 岩槻 邦男, 柏谷 博之, 田村 道夫, 堀田 満, 三浦 宏一郎 & 山岸 高旺 (1983). 植物系統分類の基礎. 北隆館. p. 20
- ^ David, J.C. (2002年). “A Preliminary Catalogue of the Names of Fungi above the Rank of Order”. Constancea, University and Jepson Herbaria. 2020年10月18日閲覧。
- ^ a b Levine, N. D., Corliss, J. O., Cox, F. E. G., Deroux, G., Grain, J., Honigberg, B. M., ... & Merinfeld, E. G. (1980). “A Newly Revised Classification of the Protozoa”. The Journal of Protozoology 27 (1): 37-58.. doi:10.1111/j.1550-7408.1980.tb04228.x.
- ^ 井上 浩, 岩槻 邦男, 柏谷 博之, 田村 道夫, 堀田 満, 三浦 宏一郎 & 山岸 高旺 (1983). 植物系統分類の基礎. 北隆館. p. 33
- ^ Perkins, F. O. (1972). “The ultrastructure of holdfasts,“rhizoids”, and “slime tracks” in thraustochytriaceous fungi and Labyrinthula spp”. Archiv für Mikrobiologie 84 (2): 95-118. doi:10.1007/BF00412431.
- ^ Alderman, D. J., Harrison, J. L., Bremer, G. B. & Jones, E. B. G. (1974). “Taxonomic revisions in the marine biflagellate fungi: the ultrastructural evidence”. Marine Biology 25: 345-357. doi:10.1007/BF00404978.
- ^ a b Tsui, C. K., Marshall, W., Yokoyama, R., Honda, D., Lippmeier, J. C., Craven, K. D., ... & Berbee, M. L. (2009). “Labyrinthulomycetes phylogeny and its implications for the evolutionary loss of chloroplasts and gain of ectoplasmic gliding”. Molecular Phylogenetics and Evolution 50 (1): 129-140. doi:10.1016/j.ympev.2008.09.027.
- ^ Patterson, D. J. (1989). “Chromophytes from a protistan perspective”. In Green, J. P. Leadbeater, B. S. C. & Diver, W. L.. The Chromophyte Algae, Problems and Perspectives. Clarendon Press. pp. 357–379. ISBN 978-0198577133
- ^ a b c Anderson, O.R. & Cavalier-Smith, T. (2012). “Ultrastructure of Diplophrys parva, a new small freshwater species, and a revised analysis of Labyrinthulea (Heterokonta)”. Acta Protozool. 51: 291–304. doi:10.4467/16890027AP.12.023.0783.
- ^ a b 巌佐庸, 倉谷滋, 斎藤成也 & 塚谷裕一 (編) (2013). “分類表”. 岩波 生物学辞典 第5版. 岩波書店. p. 1653. ISBN 978-4000803144
- ^ a b 白山 義久 (編) (2000). “分類表”. バイオディバーシティ・シリーズ (5) 無脊椎動物の多様性と系統. 裳華房. p. 265. ISBN 978-4785358273
- ^ a b 杉山 純多 (編), ed (2005). “分類表”. バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統. 裳華房. p. 397. ISBN 978-4785358273
- ^ a b “The MycoBank engine and related databases”. Robert, V., Stegehuis, G. & Stalpers, J.. 2020年10月25日閲覧。
- ^ a b Cavalier-Smith, T. (2018). “Kingdom Chromista and its eight phyla: a new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences”. Protoplasma 255: 297–357. doi:10.1007/s00709-017-1147-3.
- ^ Thakur, R., Shiratori, T. & Ishida, K. I. (2019). “Taxon-rich multigene phylogenetic analyses resolve the phylogenetic relationship among deep-branching stramenopiles”. Protist 170 (5): 125682. doi:10.1016/j.protis.2019.125682.
- ^ Cavalier-Smith, T. (1997). “Sagenista and Bigyra, two phyla of heterotrophic heterokont chromists”. Archiv für Protistenkunde 148 (3): 253-267. doi:10.1016/S0003-9365(97)80006-1.
- ^ a b Adl, S. M., Bass, D., Lane, C. E., Lukeš, J., Schoch, C. L., Smirnov, A., ... & Cárdenas, P. (2019). “Revisions to the classification, nomenclature, and diversity of eukaryotes”. Journal of Eukaryotic Microbiology 66 (1): 4-119. doi:10.1111/jeu.12691.
- ^ a b Yokoyama, R. & Honda, D. (2007). “Taxonomic rearrangement of the genus Schizochytrium sensu lato based on morphology, chemotaxonomic characteristics, and 18S rRNA gene phylogeny (Thraustochytriaceae, Labyrinthulomycetes): emendation for Schizochytrium and erection of Aurantiochytrium and Oblongichytrium gen. nov”. Mycoscience 48: 199–211. doi:10.1007/s10267-006-0362-0.
- ^ a b Yokoyama, R., Salleh, B. & Honda, D. (2007). “Taxonomic rearrangement of the genus Ulkenia sensu lato based on morphology, chemotaxonomical characteristics, and 18S rRNA gene phylogeny (Thraustochytriaceae, Labyrinthulomycetes): Emendation for Ulkenia and erection of Botryochytrium, Parietichytrium, Sicyoidochytrium gen. nov.”. Mycoscience 48: 329–341. doi:10.1007/s10267-007-0377-1.
- ^ Doi, K. & Honda, D. (2017). “Proposal of Monorhizochytrium globosum gen. nov., comb. nov.(Stramenopiles, Labyrinthulomycetes) for former Thraustochytrium globosum based on morphological features and phylogenetic relationships”. Phycological Research 65 (3): 188-201. doi:10.1111/pre.12175.
- ^ a b Takahashi, Y., Yoshida, M., Inouye, I. & Watanabe, M. M. (2016). “Fibrophrys columna gen. nov., sp. nov: A member of the family Amphifilidae”. European Journal of Protistology 56: 41-50. doi:10.1016/j.ejop.2016.06.003.
- ^ a b Pan, J., del Campo, J. & Keeling, P. J. (2017). “Reference tree and environmental sequence diversity of Labyrinthulomycetes”. Journal of Eukaryotic Microbiology 64: 88–96. doi:10.1111/jeu.12342.
- ^ a b c d Wijayawardene, N. N., Hyde, K. D., Al-Ani, L. K. T., Tedersoo, L., Haelewaters, D., Rajeshkumar, K. C. ... & Tokarev, Y. S. (2020). “Outline of Fungi and fungus-like taxa”. Mycosphere 11 (1): 1060-1456. doi:10.5943/mycosphere/11/1/8.
- ^ a b FioRito, R., Leander, C. & Leander, B. (2016). “Characterization of three novel species of Labyrinthulomycota isolated from ochre sea stars (Pisaster ochraceus)”. Marine Biology 163 (8): 170. doi:10.1007/s00227-016-2944-5.