
木部
木部 (もくぶ、英: xylem) とは、維管束植物 (シダ植物や種子植物) の維管束を構成する2つの複合組織のうちの1つであり (もう1つは師部)、道管や仮道管組織、木部繊維組織、木部柔組織からなる (図1a, b)。また、形成される場所によって、頂端分裂組織に由来する一次木部 (図1a) と、維管束形成層に由来する二次木部 (図1b) に分けられる。木本植物 (いわゆる木) では、ふつう茎や根の大部分は二次木部 (材) で占められる。木部は、水や無機養分の通道 (図1c)、および植物体の機械的支持の役割を担っている。
.jpg)
.jpg)

構成
[編集]木部は、管状要素(道管要素や仮道管)、木部繊維、木部柔細胞からなる[1][2][3](図1a, b)。これらの細胞は、木部要素 (xylem elements) ともよばれる[2]。木部柔細胞以外は、ふつう木化(リグニン化)した厚い二次細胞壁をもつ死んだ細胞(成熟した段階では原形質を失う)である。
管状要素
[編集]管状要素(かんじょうようそ、tracheary element)は管状の死細胞(成熟した状態では原形質を失う)であり、リグニンを含む二次細胞壁によって肥厚した細胞壁からなる[2][3]。管状要素の二次肥厚の様式には多様性があり、環紋肥厚、らせん紋肥厚、階紋肥厚、網紋肥厚、孔紋肥厚などがある[2][3](→詳細は道管#管状要素の肥厚様式を参照)。維管束植物の管状要素には、道管要素と仮道管がある[2][3][4]。いずれも、地下で根によって吸収された水や無機養分が地上部の茎や葉に供給される際の通路となる(→詳細は道管#木部輸送を参照)。仮道管の場合は、通道に加えて植物体の機械的支持も担っている[5][6][7]。
被子植物の多くは道管要素 (導管要素、vessel element) をもち、上下端で縦につながって道管 (導管、vessel) を形成している[2][3][6][8][9] (図2a)。道管要素どうしの隔壁には穿孔 (せん孔、perforation) とよばれる孔があり、上下の空間は連続している[3][8][10]。道管要素は仮道管(下記)にくらべると太く短いことが多い[6]。
.jpg)

被子植物以外の維管束植物(シダ植物や裸子植物)はふつう道管をもたず、仮道管(仮導管、tracheid)からなる仮道管組織(仮導管組織、tracheid tissue)が存在する[2][3][5][6](図2b)。仮道管は道管要素に較べて細長く、両端が尖った紡錘形であり、穿孔をもたない[2][3][5][6][11]。水や無機養分は細胞壁が肥厚していない部分を通して通道する[2][3][11]。この細胞壁が薄い部分はしばしば孔状であり、壁孔 (pit) とよばれる(完全な孔ではなく一次細胞壁で仕切られている)[2][3]。壁孔は道管要素にも存在する[12]。
被子植物の多くは道管をもつが、アンボレラ科、シキミモドキ科、ヤマグルマ科、さまざまな水生植物(スイレン科、ウキクサなど)のように道管をもたないものもおり、無道管被子植物とよばれる[3][6][13]。一方、シダ植物や裸子植物の中には、イワヒバ属(ヒカゲノカズラ綱)、トクサ属(トクサ亜綱)、ワラビ(ウラボシ亜綱)、グネツム類のように道管をもつものも少数知られる[2][3][6]。
木部繊維
[編集].jpg)
道管をもつ植物は、木部繊維(もくぶせんい、xylem fiber, wood fiber)からなる木部繊維組織 (xylem fiber tissue, wood fiber tissue) をもつ[1][2][3][6][14](図3)。木部繊維は細長い死細胞(成熟した状態では細胞壁のみ)であり、仮導管(上記参照)に類似しているが、より厚く肥厚してふつう通道能はほとんどなく、壁孔もあまり発達していない[1][2][6][14][15]。ただし仮道管と繊維は、下記のように中間的なものもあり、明瞭には区別できない[15]。管状要素として仮道管が主である植物では仮道管が植物体の機械的支持に用いられるが、道管が主である植物では木部繊維組織が植物体の機械的支持に用いられている[1][6][7][14]。
木部繊維はいくつかのタイプに類別されることがある。繊維仮道管[5][14](繊維状仮道管[15] fiber tracheid)は疎ではあるが有縁壁孔をもち、仮道管に似ている[3][15](上記のように仮道管と繊維の明確な区分は不可能であると考えられている[15])。一方、真正木繊維[16](師部様繊維[14] libriform wood fiber)は有縁壁孔を欠き、単壁孔をもつ[15]。また二次木部には、二次細胞壁形成後に細胞分裂して隔壁を形成した繊維が存在することがあり、隔壁繊維(隔壁細胞[3] septate fiber; 隔壁木繊維 septal wood fiber)とよばれる[14][15]。同様に原形質を残したものとして、木部柔細胞(下記)に似た代用繊維 (substitute fiber) とよばれるものもある[14]。隔壁繊維や代用繊維は機械的支持と共に養分貯蔵にも機能していると考えられている[14]。
木部柔細胞
[編集].jpg)
.jpg)
木部に存在する柔細胞は木部柔細胞(もくぶじゅうさいぼう、xylem parenchymatous cell)とよばれ、柔組織である木部柔組織 (xylem parenchyma) を形成している[1][3]。木部柔細胞は、基本的に木部における唯一の生細胞である。柔細胞はふつう薄い一次細胞壁をもつが、木部柔細胞の細胞壁はときに肥厚し、単壁孔をもつものもある[1][3]。木部柔細胞の主な機能は糖や樹脂、結晶などの貯蔵であり、ほかにも物質生産や物質輸送にも関わっている[1][3][17][18]。
二次木部(下記)の木部柔細胞には、多くの管状要素と同様に長軸方向に連なる軸方向柔組織(axial parenchyma; 紡錘形柔組織 fusiform parenchyma)と、放射方向にならんだ放射柔組織 (ray parenchyma, radial parenchyma) がある[1][3][17][18]。被子植物の軸方向柔組織は、道管とは独立して存在する独立柔組織 (apotracheal parenchyma) と、道管に接して存在する随伴柔組織 (paratracheal parenchyma) に大別される[18][19]。放射柔組織の中には、縦に1列に並んだもの(単列放射組織 uniseriate ray)や、複数列に並んだもの(多列放射組織 multiseriate ray)がある[18][19] (図4)。また放射柔細胞は、放射方向に長い平伏細胞 (procumbent ray cell)、長軸方向に長い直立細胞 (upright ray cell)、等径的な方形細胞 (square ray cell) に類別される[18]。このうち1種類の細胞だけからなるものを同形放射組織 (homocellular ray)、複数の種類の細胞が混在するものを異形放射組織 (heterocellular ray) とよぶ[18][19]。放射組織の分布様式に基づく区分もあり、同サイズの放射組織が均等に散在しているものは散在放射組織 (diffuse ray) とよばれる[18]。一方、小型の放射組織が密集しているものは集合放射組織 (aggregate ray) とよばれ(ハンノキ属など)、さらに密集化して1個の大きな放射組織となったものは広放射組織(broad ray; 複合放射組織 compound ray)とよばれる[18](コナラ属など; 図4b)。
木部柔細胞の中には、それぞれ特殊な物質を含む異形細胞(周囲の細胞と極端に異なる形をした細胞)として油細胞 (oil cell)、結晶細胞 (crystalliferous cell)、多室結晶細胞 (chambered crystalliferous cell) などが存在することがある[18]。例えばイチョウ(イチョウ科)の木部には、シュウ酸カルシウムの結晶を含む異形細胞が存在する[17]。また二次木部では、軸方向または水平方向に分泌道 (secretory canal) がしばしば見られ、乳管 (latex tube) やタンニン管 (tanniferous tube) が存在することもある[19]。球果類の木部では、エピセリウム細胞 (epithelial cell) とよばれる柔細胞で囲まれた細胞間隙が存在することがある[17]。この間隙はエピセリウム細胞が分泌する樹脂で満たされ、樹脂道 (resin canal) とよばれる(図4a)。
一次木部と二次木部
[編集]木部は、形成される場所に応じて一次木部と二次木部に分けられる[1][3][20][21]。
一次木部
[編集].jpg)
頂端分裂組織(シュート頂分裂組織、根端分裂組織)に由来する前形成層 (procambium) から形成される木部は、一次木部 (primary xylem) とよばれる[3][20]。全ての維管束植物は、一次木部をもつ。
一次木部のうち、最初に分化する部分を原生木部 (protoxylem)、その後に分化する部分を後生木部 (metaxylem) とよぶ[3][20](図5)。原生木部の管状要素は直径が小さく、二次細胞壁の肥厚様式はふつう環紋やらせん紋である[2][3][20]。一方、後生木部の管状要素は大きく発達しており、二次細胞壁の肥厚様式は階紋、網紋、または孔紋であることが多い[3][20]。原生木部は、軸の伸長や後生木部の発達に伴って崩壊し、破生細胞間隙(細胞の崩壊によって生じた細胞間隙)となることがあり、特に原生木部間隙(原生木部腔 protoxylem cavity, protoxylem lacuna)ともよばれる[3][22][23](例:スギナやススキ)。
原生木部と後生木部の位置関係(つまり木部の発生順序)は植物群およびその器官によって異なっており、以下のように類別される[20][24]。
- 外原型木部 (exarch xylem): 外端に原生木部が形成され、その後に内側の後生木部が形成される(つまり発生は求心的)。大葉植物 (大葉シダ植物と種子植物) の根(下図6a)、小葉植物の茎に見られる。
- 中原型木部 (mesarch xylem): 中間部に原生木部が形成され、その後に内側と外側に後生木部が形成される。大葉シダ植物の茎に多く見られる(下図6b)。
- 内原型木部 (endarch xylem): 内端に原生木部が形成され、その後に外側の後生木部が形成される(つまり発生は遠心的)。種子植物の茎に見られる(下図6c)。同じく遠心的に形成されるが、維管束が中心に1個だけ存在し中心の原生木部から外側へ成熟するものは特に心原型木部 (centrarch xylem) ともよばれ、リニア属などの初期維管束植物の茎、小葉植物の根に見られる[25]。
.jpg)

.jpg)
二次木部
[編集]茎や根において、一次木部と一次師部の間に生じた分裂組織である維管束形成層(単に形成層とよばれることも多い)から形成された木部は、二次木部(次生木部、二期木部、secondary xylem)とよばれる[3][21][26](下図7)。二次木部は、ふつう維管束形成層の内側に付加されていく[21](下図7)。維管束形成層による二次成長を行う植物(いわゆる木本植物) は、二次木部をもち、このような植物ではふつう茎や根のほとんどは二次木部で占められている。生物学における材、木材 (wood) は、二次木部のことを意味する[27][28]。二次木部では、一次木部にくらべて構成要素が放射方向に整然と並んでいる傾向がある[21]。


.jpg)
維管束形成層には、細長い紡錘形始原細胞 (fusiform initial) と、ほぼ等径の放射組織始原細胞 (ray initial) が存在する[21]。紡錘形始原細胞からは、道管要素や仮道管のような管状要素、木部繊維、および軸方向柔細胞が形成される[26][29]。一方、放射組織始原細胞からは、放射柔細胞が形成され、球果類では放射方向に伸びる放射仮道管 (ray tracheid) も形成されることがある[17][29][30]。また被子植物では、2つの道管要素を穿孔を通してつなぐ特殊な細胞が形成されることがあり、有穿孔放射組織細胞 (perforated ray cell) とよばれる[18][19]。このような放射組織始原細胞に由来する細胞からなる組織は、放射組織 (ray) とよばれる[3][17][30](上図7)。同一の放射組織始原細胞に由来する放射組織は、木部へ伸びるもの (木部放射組織、xylem ray) と、師部へ伸びるもの (師部放射組織、phloem ray) が連続している[30](上図7c)。放射組織の機能は、同化産物など物質の貯蔵と、放射方向の物質輸送にある[18]。
心材と辺材
[編集]

軸(茎や根)の周縁部にある二次木部、つまりより新しい二次木部は辺材(splint wood、液材 sapwood)とよばれる[27][28][31][32]。辺材は軸方向柔組織や放射柔組織など生きた細胞を含み、支持機能と共に通道機能や貯蔵機能をもつ[28][32]。色素などをあまり含んでいないため、心材にくらべて色が薄く、俗に白太、白材ともよばれる[28][32](図8)。一方、軸の中心部にある二次木部、つまりより古い二次木部は心材 (heartwood, heart wood) とよばれる[27][31][33]。心材には生きた細胞が全く存在せず、通道機能や貯蔵機能が失われている[33][31]。一方でふつう硬化し、機械的支持機能が増している[28][33]。リグニンやポリフェノールなどが多く沈着し、しばしば着色しているため、俗に赤心、赤身、赤味、赤肌、赤材ともよばれる[28][33](図8)。コクタン(カキノキ科)やシタン(マメ科)はその名が示す色をした心材が利用されている[28](図8b)。またロッグウッド(マメ科)の心材から抽出されるヘマトキシリンは、細胞核などの染色剤として用いられる[28]。辺材と心材の境界部は移行材 (intermediate wood, transition zone) とよばれ[32][34]、フラボノイド合成系などの活性が高く、ポリフェノールやフラボノイドなどの心材成分が生成されている[31]。一方で、トドマツ(マツ科)やアオダモ(モクセイ科)のように心材成分が少なく、心材と辺材の区分が不明瞭なものもいる[27]。
また辺材でも、その全てが通水に用いられているわけではない。最も新しい(つまり最外の)年輪のみで通水するもの(例: ヤマウルシ)、各年輪の外側(後半部)で通水するもの(例: ネコヤナギ)、各年輪の内側(前半部)で通水するもの(例: イヌブナ)などが知られている[8]。
年輪
[編集].jpg)
.jpg)
維管束形成層から1年間に形成された二次木部は管状になり、年輪 (annual ring) とよばれる[26][28][35]。四季がある地域では維管束形成層の活動が季節によって大きく変動するため、1年ごとの年輪が明瞭に区別できることが多い(上図9)。春から初夏にかけてつくられた二次木部は早材 (early wood, earlywood; 春材 spring wood) とよばれ、管状要素(道管要素や仮道管)の径が太く、比較的柔らかく明色であることが多い[27][28][32]。一方、夏から秋にかけてつくられた二次木部は晩材 (late wood, latewood; 夏材 summer wood; 秋材 autumn wood) とよばれ、管状要素の径が細く緻密であり、比較的硬く色が濃いことが多い[27][28]。晩材と次の年の早材の間には明瞭な境界が存在することが多く、この境界は年輪界 (annual ring boundary) とよばれる[28][35](上図9b)。一方、四季が不明瞭な地域(熱帯)では、年輪がはっきりしないことが多い。また雨期と乾期に対応して形成された年輪様の構造は成長輪 (growth ring) ともよばれる[35]。年輪の幅は環境条件によって変動するため、これをマーカーとした木材の年代推定が広く行われている(年輪年代学[注釈 1])。
材のタイプ
[編集]
二次木部(材)は、道管の有無やその配列などに多様性があり(図10)、それに応じて以下のように類別される[8][28][37][38][39]。材における各要素の形、大きさ、密度、配列様式などには大きな多様性があり、材から樹種を同定できる[19][40]。
- 無孔材 (nonporous wood, non-pored wood)(図10下、下図11a)
- 有孔材 (porous wood, pored wood)(図10上、下図11b–e)
- 道管をもつ材。木部繊維をもつため一般的に針葉樹材(無孔材)よりも硬く、商業的慣習から硬材 (hardwood) ともよばれる[41][42][43]。ただしバルサ(アオイ科)のように柔組織を多く含むため非常に柔らかい材をもつ種もいる[19]。
- 散孔材 (diffuse-porous wood)(下図11b)
- 半散孔材 (semidiffuse-porous wood、半環孔材)(下図11c)
- 環孔材 (ring-porous wood)(下図11d, e)
_(20623135529).jpg)
_(20623120059).jpg)
_(20800419382).jpg)
_(20783581546).jpg)
_(20816905591).jpg)
また横断面(木口面)からみた道管の配列様式(環孔材の場合は特に晩材部において)に応じて以下のように類別されることもある[8][28][44]。
- 放射状 (radial pattern): 道管が放射方向にならぶ。シラカシ(ブナ科)、アカシデ(カバノキ科)、カキノキ(カキノキ科)など。
- 接線状 (tangentional pattern): 道管が接線方向にならぶ。ハリギリ (ウコギ科) など。放射状と接線状の中間的なもの(斜めにならぶ)は斜線状 (diagonal pattern) とよばれる。
- 火炎状(紋様状)(dendritic pattern): 道管が紋様状に集まって無孔領域と明瞭に分かれる。雑孔材 (mosaic-porous wood)、紋様孔材 (figured-porous wood) ともよばれる。ヒイラギ、モクセイ(モクセイ科)、ゴヨウツツジ(ツツジ科)など。
ギャラリー
[編集]-

-

-

-
-

-

-

-

-
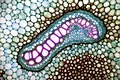
 マツ属(マツ科)の2年目の茎の横断面
マツ属(マツ科)の2年目の茎の横断面 -
 マツ属(マツ科)の二次師部(左)と維管束形成層、二次木部の横断面
マツ属(マツ科)の二次師部(左)と維管束形成層、二次木部の横断面 -
 ニワトコ属(ガマズミ科)の二次師部(上)と維管束形成層、二次木部の横断面であり、放射柔組織も見える。
ニワトコ属(ガマズミ科)の二次師部(上)と維管束形成層、二次木部の横断面であり、放射柔組織も見える。 -
 ユリノキ属(モクレン科)の木部縦断面: 道管の穿孔が見える。
ユリノキ属(モクレン科)の木部縦断面: 道管の穿孔が見える。 -
 マツ属(マツ科)の二次木部(仮道管)横断面
マツ属(マツ科)の二次木部(仮道管)横断面 -
 マツ属(マツ科)の二次木部横断面であり、放射柔組織が見える。
マツ属(マツ科)の二次木部横断面であり、放射柔組織が見える。 -
 マツ属(マツ科)の二次木部横断面であり、樹脂道と放射柔組織が見える。
マツ属(マツ科)の二次木部横断面であり、樹脂道と放射柔組織が見える。 -
 マツ属(マツ科)の二次木部(仮道管)縦断面 (放射断面)
マツ属(マツ科)の二次木部(仮道管)縦断面 (放射断面) -
 マツ属(マツ科)の二次木部縦断面 (放射断面)
マツ属(マツ科)の二次木部縦断面 (放射断面) -
 マツ属(マツ科)の二次木部縦断面(接線断面)
マツ属(マツ科)の二次木部縦断面(接線断面) -

-

-
 マンソニア(アオイ科)の茎の二次木部の横断面
マンソニア(アオイ科)の茎の二次木部の横断面 -
 マンソニア(アオイ科)の茎の二次木部の縦断面(接線面)
マンソニア(アオイ科)の茎の二次木部の縦断面(接線面) -

-
 ナラ(ブナ科)材断面の走査型電子顕微鏡像(上)と光学顕微鏡像(下)
ナラ(ブナ科)材断面の走査型電子顕微鏡像(上)と光学顕微鏡像(下)
.jpg)
.jpg)
.jpg)

.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)
.jpg)



.jpg)

脚注
[編集]注釈
[編集]出典
[編集]- ^ a b c d e f g h i 『岩波 生物学辞典 第5版』 (2013) 「木部」 p.1396
- ^ a b c d e f g h i j k l m n 原 (1994) pp.80–85, 91
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa 清水 (2001) 「木部」 pp.182–185
- ^ 『岩波 生物学辞典 第5版』 (2013) 「管状要素」 p.264
- ^ a b c d 『岩波 生物学辞典 第5版』 (2013) 「仮道管」 p.232
- ^ a b c d e f g h i j 岡本 (1997) pp.134–138
- ^ a b 大山 (2011) pp.21–25
- ^ a b c d e 佐野 & 内海 (2011) pp.59–66
- ^ 『岩波 生物学辞典 第5版』 (2013) 「道管」 pp.977–978
- ^ 『岩波 生物学辞典 第5版』 (2013) 「穿孔」 pp.803–804
- ^ a b 佐野 (2011) pp.53–55
- ^ 佐野 (2011) pp.53–55
- ^ 田村 (1999) pp.137-143
- ^ a b c d e f g h 『岩波 生物学辞典 第5版』(2013) 「繊維」 p.799
- ^ a b c d e f g 佐野 & 内海 (2011) pp.67–68
- ^ 『学術用語集 植物学編』 (1990) p.230
- ^ a b c d e f 佐野 (2011) pp.55–58
- ^ a b c d e f g h i j k 佐野 & 内海 (2011) pp.68–73
- ^ a b c d e f g ルダル (1997) pp.41–52
- ^ a b c d e f 『岩波 生物学辞典 第5版』(2013) 「一次木部」 p.73
- ^ a b c d e 『岩波 生物学辞典 第5版』(2013) 「二次木部」 p.1036
- ^ 高部 (2011) pp.77–78
- ^ 『学術用語集 植物学編』 (1990) p.47
- ^ ギフォード & フォスター (2002) pp.48–51
- ^ Simpson (2006) p.78
- ^ a b c 原 (1994) pp.134–139
- ^ a b c d e f 船田 (2011) pp.15–20
- ^ a b c d e f g h i j k l m n 清水 (2001) 「材」 pp.193–195
- ^ a b 船田 (2011) pp.46–52
- ^ a b c 『岩波 生物学辞典 第5版』 (2013) 「放射仮道管」 p.1296, 「放射組織」 p.1299
- ^ a b c d 中田 & 船田 (2011) pp.117–124
- ^ a b c d e 『岩波 生物学辞典 第5版』(2013) 「辺材」 p.1285
- ^ a b c d 『岩波 生物学辞典 第5版』(2013) 「心材」 p.703
- ^ 中田 (2014) pp.63–79
- ^ a b c 『岩波 生物学辞典 第5版』(2013) 「年輪」 p.1061
- ^ 『考古学キーワード』 (1997) pp.58–59
- ^ 『岩波 生物学辞典 第5版』 (2013) 「材」 p.507
- ^ 『岩波 生物学辞典 第5版』(2013) 「散孔材」 p.550
- ^ 『岩波 生物学辞典 第5版』(2013) 「環孔材」 pp.260–261
- ^ 木材データベース. 森林総合研究所. (2020年4月21日閲覧)
- ^ a b 清水 (2001) pp.23–24
- ^ a b 『岩波 生物学辞典 第5版』 (2013) 「無孔材」 pp.1368–1369
- ^ a b Bowes & Mauseth (2008) pp.68–71
- ^ 『岩波 生物学辞典 第5版』 (2013) 「紋様孔材」 p.1402
参考文献
[編集]一般書籍
[編集]- 福島和彦・船田良・杉山淳司・高部圭司・梅澤俊明・山本浩之 編『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年10月、593頁。ISBN 978-4860992521。
- 船田良「木材の構造と形成」『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年、15-20頁。
- 大山幹成「木材の構造と進化」『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年、21-25頁。
- 船田良「木部細胞の分化・成熟」『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年、46-52頁。
- 佐野雄三「仮道管」『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年、53-55頁。
- 佐野雄三「軸方向柔細胞、放射組織、樹脂道とエピセリウム細胞」『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年、55-58頁。
- 佐野雄三 & 内海泰弘「道管」『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年、59-66頁。
- 佐野雄三 & 内海泰弘「木部繊維」『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年、67-68頁。
- 佐野雄三 & 内海泰弘「軸方向柔細胞、放射組織」『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年、68-73頁。
- 高部圭司「単子葉植物の構造と形成 道管」『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年、77-78頁。
- 佐野雄三「壁孔」『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年、93-97頁。
- 中田了五 & 船田良「心材の構造と形成」『木質の形成 第2版 -バイオマス科学への招待-』海青社、2011年、117-124頁。
- 安蒜政雄 編『考古学キーワード』有斐閣〈有斐閣双書〉、1997年11月、252頁。ISBN 4-641-05860-1。
- 岡本素治 著「9章 被子植物にみる多様性と系統」、加藤雅啓 編『植物の多様性と系統』裳華房〈バイオディバーシティ・シリーズ (2)〉、1997年10月、131-180頁。ISBN 978-4-7853-5825-9。
- 田村道夫『植物の系統』文一総合出版、1999年2月、222頁。ISBN 978-4829921265。
- 原襄『植物形態学』朝倉書店、1994年7月、180頁。ISBN 978-4254170863。
- 清水建美『図説 植物用語事典』八坂書房、2001年7月、323頁。ISBN 978-4896944792。
- アーネスト・ギフォード & エイドリアンス・フォスター 著、長谷部光泰・鈴木武・植田邦彦 訳『維管束植物の形態と進化』文一総合出版、2002年3月、643頁。ISBN 978-4829921609。
- ポーラ・ルダル 著、鈴木三男・田川裕美 訳『植物解剖学入門 ―植物体の構造とその形成―』八坂書房、1997年4月、197頁。ISBN 978-4896946963。* Bowes, B. & Mauseth, J. D. (2008). “Xylem, Structure of wood”. Plant Structure: A Colour Guide 2nd Edition. Jones & Bartlett Learning. pp. 288. ISBN 978-0763763862
- Simpson, M. (2006). Plant Systematics. Academic Press. pp. 590. ISBN 978-0126444605
事典・辞典等
[編集]- 巌佐庸、倉谷, 滋、斎藤, 成也 ほか 編『岩波 生物学辞典 第5版』岩波書店、2013年2月、2192頁。ISBN 978-4000803144。
- 文部省、日本植物学会 編『学術用語集 植物学編 (増訂版)』丸善、1990年3月、684頁。ISBN 978-4621035344。
雑誌論文
[編集]- 中田了五「樹木の wetwood 現象と定義」『木材学会誌』第60巻、2014年、63-79頁、NAID 130004775936。
関連項目
[編集]外部リンク
[編集]- 福原達人 (2019) 1-3. 木部・篩部. 植物形態学. 福岡教育大学. (2020年3月7日閲覧)
- 維管束 光合成事典. 日本光合成学会. (2020年3月7日閲覧)
- 木材データベース. 森林総合研究所. (2020年3月25日閲覧)
- 東南アジア産材の木材特性データベース. 森林総合研究所. (2020年3月25日閲覧)
- 『木部』 - コトバンク