
ヒヨケムシ
| ヒヨケムシ | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 様々なヒヨケムシ[注釈 2]
| ||||||||||||||||||
| 地質時代 | ||||||||||||||||||
| 古生代石炭紀ペンシルベニア紀(約3億600万年前)[注釈 1][1] - 現世 | ||||||||||||||||||
| 分類 | ||||||||||||||||||
| ||||||||||||||||||
| 学名 | ||||||||||||||||||
| Solifugae Sundevall, 1833 [2] | ||||||||||||||||||
| シノニム | ||||||||||||||||||
| ||||||||||||||||||
| 和名 | ||||||||||||||||||
| ヒヨケムシ | ||||||||||||||||||
| 英名 | ||||||||||||||||||
| Solifugid [4] Solifuge [4] Solpugid [4] Camel spider [4] Sun spider [4] Sun scorpion [4] Wind spider [4] Wind scorpion [4] Red roman [4] Roman spider [5] Jerrymander [6] Jerrymunglum [6] Kalahari Ferrari [7] Child of the Earth [8] | ||||||||||||||||||
| 科 [4] | ||||||||||||||||||
ヒヨケムシ(ヒヨケムシ類、学名: Solifugae[2])は、鋏角亜門クモガタ綱に分類される節足動物の分類群の一つ。分類学上はヒヨケムシ目とされる。強大なはさみ型の鋏角と脚のような触肢を先頭に有し、主に砂漠に生息する俊敏な捕食者である[9][10][6][4]。1,100種以上知られ、最古の化石記録は約3億年前の石炭紀まで遡る[1][11]。
一見クモにも似た姿だが、クモではない。研究は少なく、その形態・生態・系統に関しては未だに不明点が多い[10][12][4][13]。
名称
[編集]学名「Solifugae」(古典的には「Solpugida」ともされる[5][9][3])はラテン語の「sol」(太陽)[14]と「fugere」(逃げる)[15]の合成語であり、あわせて「太陽から逃げる者」を意味する[6]。これは日当たりを避け、巣穴や日当りのない場所に身を隠す習性に由来する[6]。学名に因んだ学術的な通称として「solifugid」「solifuge」「solpugid[注釈 3]」が挙げられる[6]。和名は「ヒヨケムシ」と呼ぶ[16]。
英語やアフリカーンス語など、原産地の言語では数多くの俗称が知られ、次の通りに挙げられる[6]。ただし、これらの多くはクモ(spider)やサソリ(scorpion)など別のクモガタ類を指す名称を含む、もしくはあらゆる不正確な情報(後述参照)に由来するものであり、誤解を招きやすい[5][17][6][18][19]。また、大まかな姿はサソリよりクモに似るため、その中で「クモ」が付くほうが常用される傾向がある[6]。なお、古代ギリシア語では明確にクモ(ἀράχνη, arachne)から区別され、脚10本(実際には触肢2本と脚8本)のような姿から「φαλάνγιον」(phalangion, "指骨")と呼ばれている[20][21]。
- Camel spider("ラクダのクモ")
- Wind spider("風のクモ"), Wind scorpion("風のサソリ"), Kalahari Ferrari("カラハリのフェラーリ")
- Sun spider("太陽のクモ"), Sun scorpion("太陽のサソリ")
- Red roman, Roman spider
- Jerrymander, Jerrymunglum
- Child of the Earth("大地の子供")[8]。
- Haarskeerder("髪を切る者"), Baardskeerder("髭を切る者")
- Jag spinnekoppe("狩りクモ")[20][6]
- Gift-kanker("毒グモ")[20][6]
- Vetvreter("脂肪を食べる者")[20][6]
形態
[編集]
A: 背面、B: 側面、C: 腹面、an: 肛門、ch: 鋏角、go: 生殖孔、l1-4: 第1-4脚、me: 中眼、ms: mesopeltidium、mt: metapeltidium、opi: 後体、p: propeltidium、pe: 触肢、pro: 前体、ra: ラケット器官、sp: 気門、st: 腹板、te: 背板
-

-
 鮮やかで昼行性の Metasolpuga picta
鮮やかで昼行性の Metasolpuga picta


本項は特記しない限り、成体の形態について記述する(幼生の形態については生活環の項を参照のこと)。
体長は種類により微小な数mm[22]から巨大な10cmに及ぶ[9]。多くは1-7cmに当たる[20]が、陸生節足動物にしては大型な種をも含め、最大級のものは附属肢を含めて16cmとなる[6]。体は前体と後体という2つの合体節からなり、その間はくびれている[23]。他のクモガタ類と同様に脚4対(8本)であるが、先頭1対の触肢は脚のように発達したため、脚5対(10本)と見間違われやすい。付属肢と先頭周辺以外の表皮(クチクラ)はほとんど柔らかく、多くは浅い黄色やクリーム色などの薄い体色をしている[20]が、赤・黒・白などの色や模様をした派手な種類もあり[4]、前者は夜行性、後者は昼行性の種類に多い[6]。全身に長さが不均一な毛(剛毛 setae)が生えて、一部の種類はそれが特に発達で、煌々たる毛に覆われた玉のような姿となる[6]。
分節した前体・強大な鋏角・発達した気管系など、鋏角類として独特な性質をもつ[23]。他のクモガタ類から明確に区別できるヒヨケムシの固有派生形質は、強大化した2節のはさみ型の鋏角・触肢先端の吸盤・第4脚のラケット器官・雄成体の鋏角に特有の鞭毛複合体などが挙げられる[10][4][23]。
前体
[編集]
暗灰色: 背甲(背板)、A: 背面、B: 側面、C: 腹面、aa: anterior arc、ap: posterior arc、ch: 鋏角、ep: 口上板、l1-4: 第1-4脚、le: anterolateral propeltidial lobe、ll: 触肢基節中央の内突起、lma: lamina exterior major、lmi: lamina exterior minor、me: 中眼、mp: median plagula、ms: mesopeltidium、mt: metapeltidium、ot: 眼丘、p: propeltidium、pe: 触肢、pen: 触肢基節両前端の内突起、ra: ラケット器官、sp: 気門、st: 腹板、v: 口腹面の外骨格
他のクモガタ類と同様、前体(prosoma, 頭胸部 cephalothorax ともいうが、頭部そのものに相当[24])は眼や口が由来する先節と、順に鋏角1対・触肢1対・脚4対という計6対の付属肢(関節肢、詳細は後述参照)が由来する第1-6体節でできた合体節である[23]。なお、前体は合体節的に頭部そのものである[24]にもかかわらず、ヒヨケムシはその後半が高度に分節したため、全体的に昆虫の頭部と胸部を彷彿とさせる[25][26]。
背面構造
[編集]

ヒヨケムシの前体背面の外骨格、いわゆる背甲(carapace, prosomal dorsal shield)は他の多くのクモガタ類(単一の甲羅で分節がない)とは異なり、分節が進み、大まかに前後で3枚の「peltidium」という遊離した背板(tergite)に分かれている[26][23]。
先節と第1-4体節(眼・鋏角・触肢・第1-2脚)に対応する前半部の背板(propeltidium)は丈夫で大きく膨らみ、鋏角の基部に繋がる筋肉を大量に詰まる[26][20]。両前端は鋏角と関節した1対の外骨格(lateral lobe, exterior lobe[5], lobus exterior[17], anterolateral propeltidial lobe[4])があり、種類により後方が propeltidium と癒合、もしくは完全な境目で propeltidium と分かれている[26]。後縁の周辺には細かな外骨格(parapeltidium)が数枚あり、順に両前方の「anterior arc」(arcus anterior)・中央の「median plagula」・両後方の「posterior arc」(arcus posterior)に分かれるが、種類により融合と発達具合が異なり、それぞれ特定の体節に対応するとも考えられる[26][23]。後半2枚の背板(mesopeltidium と metapeltidium)は柔らかく、第5-6体節(第3-4脚)に対応する[23][注釈 5]。このように第4-5体節の間から分節し始めた前体は、コヨリムシ・胸板ダニ類(Acariformes)・ヤイトムシにも見られる[27][23]。
背甲の前端中央はやや盛り上がった眼丘(ocular tubercles)で、中眼(median eye)由来の単眼が1対ある。なお、一部の種類は、鋏角と関節した両前端に痕跡的な側眼(lateral eye)と思われる構造も1対以上兼ね備えている[26][28][29]。
腹面構造
[編集]
chel: 鋏角、cx pp: 触肢基節、cxI-IV: 第1-4脚基節、st: 腹板、矢印: 第2-3脚基節の境目
前体の腹面はほぼ完全に触肢と第1-4脚(後述)の付け根(基節 coxa)に占められ、一見して腹板(sternite)をもたないが、実際、第1脚基節の間には目立たない腹板(tritosternum)を1枚もつ[26][5]。これらの外骨格は癒合が進んで前後2枚の大きな板となり、触肢基節・第1-2脚基節・腹板が前の板、第3-4脚基節が後ろの板を構成する[30]。この2枚の板の境目、いわゆる第2-3脚基節の間の節間膜には、スリット状の構造体に覆われた1対の気門(spiracle, stigma)が開く[26][17][31]。
口など
[編集]).jpg)
)_-_21079857215.jpg)
口は鋏角と触肢の間で開口し、咽頭の背面が縦長い口上板(epistome, 口上突起とも、口上板と上唇 labrum の複合体 epistomo-labral plate ともされる[32])、両腹面が触肢基節由来の1対の内突起(lateral lip, mouth lobe[注釈 6])[33][32]、腹面中央が1枚の由来不明な外骨格(ventromedian sclerite)[注釈 7]に覆われている[34]。これらの構造は普段では鋏角に覆われて目立たなく、お互いに癒合して前向きの嘴状の複合体(rostrum)をなしている[26][32][34]。口上板と触肢基節内突起の先端には、それぞれブラシ状と鞭状の剛毛が生えている[26][20][32]。
鋏角
[編集]
).jpg)
).jpg)
ヒヨケムシの最も目立つ特徴は、前体の正面から突き出した強大な鋏角(chelicerae)である。往々にして前体の半分以上を占めるほど長く、クモガタ類の中で最大の鋏角をもつとも言われている[4]。2節の肢節に構成され、大きな鋏となっている。基部側の肢節(paturon)は大量の筋肉が詰まった肥厚な掌部(manus)で、付け根の側面が背甲の両前端に関節する支点となる[26][32][4]。前背側に不動指が突出し、前腹面に関節する先端側の肢節が可動指で上下に動く[26][4]。両指の内側(不動指の腹側と可動指の背側)には大小の歯、掌部の内側には数多くの剛毛が並び、カルシュヒヨケムシ科以外では摩擦音を出す用の構造(stridulatory apparatus)もある[35][4]。鋏角の特徴、特に歯と雄の鞭毛複合体(詳細は後述参照)の形は種の同定形質として重要視されている[4]。また、外見上では左右対称だが、噛む力にある程度の非対称性をもつ種類もある[36]。
触肢と脚
[編集]).jpg)

-

-
 Zeria venator の吸盤背面
Zeria venator の吸盤背面 -
 Zeria venator の吸盤腹面
Zeria venator の吸盤腹面
).jpg)
)_Abb_76_B.jpg)
)_Abb_76_A.jpg)
).jpg)
脚5対10本に見えるが、他のクモガタ類と同じく4対8本で、最初1対の長大な付属肢は脚ではなく触肢(pedipalp)である。触肢は見かけ上6節に分れ[注釈 8]、基節は左右癒合した不動な構造体で、前に突出して鋏角の付け根腹面を覆いかぶさり、1対の指状の内突起(endite)を両前端に、口の両腹面を覆いかぶさった細い内突起(前述参照)を中央にもつ[26][32][34]。第3-5肢節は長大で、短い第6肢節末端の前跗節(pretarsus, apotele[5])に爪はなく、代わりに特殊な吸盤(suctorial organ)がある。なお、この吸盤は普段では内部に格納され、上下2枚の蓋に覆われている[37][38][39]。
).jpg)
).jpg)
4対の脚は前体の両腹面から左右に張り出している。見かけ上統一に(末端の目立たない前跗節と二次的な跗小節を除いて)7節でできた他の多くのクモガタ類の脚とは異なり、ヒヨケムシの脚の見かけ上の肢節数は変則的で、第1-2脚は7節、第3-4脚は8節に分れている[26][注釈 8][40]。基節は不動で、第1-2脚基節は触肢基節と腹板に、第3-4脚基節はお互いに癒合する[30]。第1-2脚の第4-5肢節と、第3-4脚の第5-6肢節の関節に伸筋はなく、代わりに弾性のある特殊な外骨格(elastic sclerite)で関節を伸ばす[40][41]。第1脚は華奢で歩行に用いられず、補助的な感覚器官となっている[40][20][17]。残り3対は歩行用の歩脚(walking leg)で、跗節は分類群や番目により二次的な分節(跗小節 tarsomere)があったりなかったりする[26][42][17]。歩脚は通常では後方ほど発達で、地表での歩行に適した形(cursorial)であるが、アナホリヒヨケムシ科の場合は地下性に適した特殊な形(fossorial)で[9][43]、第4脚が第3脚より短い[26]。第4脚の第1-3肢節腹面には、5本(ほとんどの種類)もしくは2-3本(アナホリヒヨケムシ科の一部)のラケット器官(malleoli, racquet organ)という扇形の感覚器官がぶら下がって並んでいる[26][20][44]。各脚末端の前跗節は原則として爪間盤(pulvillus, arolium, empodium, onychium, plantium[5])と1対の爪(tarsal claw)を有し、爪は途中に分節がある[26][17]。ただし第1脚の爪間盤と爪は退化的で、種類により消失した場合もある[26][42][17]。第2-3脚の脛節(第2脚の第5肢節、第3脚の第6肢節)外側には、砂を掘る用の強大な棘が並んでいる[45]。
肢節の相同性
[編集]-

-
 ヒヨケムシの第3-4脚の転節と脛節の間の関節構造
ヒヨケムシの第3-4脚の転節と脛節の間の関節構造


カニムシに似て、ヒヨケムシの触肢と脚は、腿節(femur)と脛節(tibia)に見える肢節の間に典型的な膝節(patella, 通常では膝に当たる所にある短い肢節)が見当たらず、脚の基部の短い肢節も他のクモガタ類より1-2節多いため、古典的には膝節をもたない同時に、第1-2脚が分化した腿節、第3-4脚が分化した転節(trochanter)と腿節をもつと解釈されてきた[26][46]。しかし筋肉の構造に基づくと、脚の腿節に見える長い肢節(第1-2脚の第4肢節、第3-4脚の第5肢節)は発達した膝節で、その直前の短い肢節(第1-2脚の第3肢節、第3-4脚の第3-4肢節)の方が腿節だと示される[40][47]。触肢の場合、古典的に脛節と蹠節(metatarsus, basitarsus)扱いされた第4-5肢節がそれぞれ膝節と脛節である(蹠節はない)[5]。第3-4脚の第2-3肢節は古典的には2節に分化した転節と解釈されきてたが、同様に筋肉の構造に基づくと、第2肢節のみが転節で、第3肢節は2節に分化した腿節の前半部だと示される[40][47][41]。また、この再検証を踏まえると、ヒヨケムシの脚の腿節-膝節関節は他のクモガタ類(原則として双関節丘のヒンジ関節)とは大きく異なり、1つの関節丘のみをもつとなる[40]。
| 肢節の番目 | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| 膝節なし説 | 基節 coxa | 転節 trochanter | 腿節 femur | 脛節 tibia | 蹠節 metatarsus / 基跗節 basitarsus | 跗節 tarsus / 端跗節 telotarsus |
| 膝節あり説 | 基節 coxa | 転節 trochanter | 腿節 femur | 膝節 patella | 脛節 tibia | 跗節 tarsus |
| 肢節の番目 | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| 膝節なし、転節2節説 | 基節 coxa | 転節 trochanter | 第1腿節 femur 1 / 前腿節 prefemur / basifemur | 第2腿節 femur 2 / 後腿節 postfemur / telofemur | 脛節 tibia | 蹠節 metatarsus / 基跗節 basitarsus | 跗節 tarsus / 端跗節 telotarsus |
| 膝節あり説 | 基節 coxa | 転節 trochanter | 腿節 femur | 膝節 patella | 脛節 tibia | 蹠節 metatarsus / 基跗節 basitarsus | 跗節 tarsus / 端跗節 telotarsus |
| 肢節の番目 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| 膝節なし、転節2節説 | 基節 coxa | 第1転節 trochanter 1 | 第2転節 trochanter 2 | 第1腿節 femur 1 / 前腿節 prefemur / basifemur | 第2腿節 femur 2 / 後腿節 postfemur / telofemur | 脛節 tibia | 蹠節 metatarsus / basitarsus | 跗節 tarsus / telotarsus |
| 膝節あり、腿節2節説 | 基節 coxa | 転節 trochanter | 第1腿節 femur 1 / 前腿節 prefemur / basifemur | 第2腿節 femur 2 / 後腿節 postfemur / telofemur | 膝節 patella | 脛節 tibia | 蹠節 metatarsus / basitarsus | 跗節 tarsus / telotarsus |
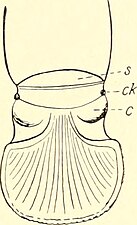
後体
[編集]
2-11: 後体第2-11節(見かけ上の第1-10節)、an: 肛門、go: 生殖孔、gop: 生殖口蓋、sp: 気門、st: 腹板、te: 背板

-

-
 左右に分かれた背板をもつ Hexisopus abnormis
左右に分かれた背板をもつ Hexisopus abnormis


後体(opisthosoma, 腹部 abdomen ともいうが、胴部に相当[24])は伸縮性をもつ縦長い楕円形で、柔らかく、上下が一連の背板と腹板[23]、左右が幅広い節間膜に覆われている[26][46]。背板と腹板は種類や番目により左右2枚に分けれた場合がある[26]。外見上は10節に見えるが、実際には11節の体節が含まれた合体節で、第1節は著しく退化して目立たない[26][24][23][注釈 5]。第2節(見かけ上の第1節)腹面は縦割れの生殖口蓋(genital operculum, genital sternite)で、生殖孔(gonopore, genital orifice)はその中央に開く[26]。5つの気門は第3-5節(見かけ上の第2-4節)のそれぞれの腹面の後縁中央に並び、そのうち前の2対は第3-4節、残り1つは第5節に開く[17][31]。この第5節の気門はヒヨケムシに特有で、元々1対だった気門から癒合したものだと考えられる[31]。前体のスリット状の気門に比べると、後体の気門は単調な穴で特に複雑な構造はない[17]。最終体節(後体第11節、見かけ上の第10節)に尾節(telson)はなく[23]、肛門はその末端(ほとんどの種類)もしくは腹面(オオヒヨケムシ科と Dinorhax 属)に開く[42][29]。
付属肢らしき構造は見当たらないが、生殖孔と気門を覆いかぶさった腹面の外骨格は腹板ではなく、四肺類(クモ・ウデムシ・サソリモドキなど)に見られるような付属肢由来の蓋板(operculum)とも解釈される[46][24]。
雌雄
[編集]



-
![雄の鋏角と鞭毛[注釈 9]](//upload.wikimedia.org/wikipedia/commons/thumb/d/dd/Solifugae_flagellum.png/289px-Solifugae_flagellum.png)
-
![雄の鋏角と setiform flagellar complex[注釈 10]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b8/Solifugae_setiform_flagellar_complex.png/378px-Solifugae_setiform_flagellar_complex.png) 雄の鋏角と setiform flagellar complex[注釈 10]
雄の鋏角と setiform flagellar complex[注釈 10]
![雄の鋏角と鞭毛[注釈 9]](/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Solifugae_flagellum.png)
![雄の鋏角と setiform flagellar complex[注釈 10]](/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Solifugae_setiform_flagellar_complex.png)
生殖口蓋は雌雄によって構造が異なり、ラケット器官は雄の方が発達する[48][13]。雄に特有の最大の特徴は、鋏角不動指の内側に生えて、特化した剛毛でできた鞭毛複合体(flagellar complex)である[4][48][13]。これは種類によって鞭状や膜状などに発達した1-2本の鞭毛(flagellum, primary flagellum)を主体にしたもの、もしくは複数の同形な剛毛でできたもの(setiform flagellar complex)である[注釈 11][4]。雄の鞭毛複合体は種類によって形態が異なり、ヒヨケムシの種特異性を表した重要な同定形質とされる[4]。
前述の性質以外でも、顕著な性的二形が見られる種類が多い。多くの場合、雄は明らかに雌より華奢で、触肢と脚も雌より長い[48]。鋏角は鞭毛複合体の有無以外では種類により性的二形の程度が異なり、雌雄ほぼ同形なものがあれば、雄の方が明らかに華奢で歯が貧弱なものもあり、雄の不動指が変わった形に特化したものもある[4]。一部の種類[注釈 12]は、雄のみ後体の気門周辺に丈夫な突起物に特化した剛毛(ctenidia)が並んでいる[49]。触肢の棘・眼丘・背板[50]などが雌雄により構造が異なる種類もある[13]。
生殖腺
[編集]

ヒヨケムシは雌雄とも後体の生殖孔内側にある1つの房室(genital chamber)から対になる生殖腺が後方に続く[17]。雌性生殖腺は1対の短い卵管(oviduct)と、大小の卵母細胞(oocyte)が密生した長大な卵巣(ovary)からなる[17]。雄性生殖腺は1対の肥厚な精管(vasa deferens)とその末端から2つに枝分かれた2対の精巣(testis)からなり、一部の種類は房室の左右に1対の嚢(lateral pouch)がある[17]。雄成体の精巣は萎縮し、成体になるまで生成された精子を精管に蓄えている[13]。精子は丸みを帯びた単調な形で、鞭毛がない[51][52][43]。これら構造は雌雄とも種類によってやや異なる場合があり[17]、例えばヒヨケムシ科の卵管は複数の分岐を有し[13]、サメヒヨケムシ科の精子は他の科より細長い[52]。
他の内部構造
[編集]).jpg)
).jpg)

).jpg)



呼吸器
[編集]ヒヨケムシの呼吸器は気管系(tracheal system)であるが、クモガタ類にしては異様に発達で、その構造は全体・細部とも昆虫のものを彷彿とさせる(収斂)[17][31]。前体2つ(1対)と後体5つ(2対+1つ)の気門に繋がる気管は全身を貫通して所々に分岐と連合があり、鋏角・触肢・脚にも及ぶ[31]。少なくとも大型種(知られる限りサメヒヨケムシ科とスナハシリヒヨケムシ科)は、鋏角内部の気管が数対の気嚢(air sac)に発達する[31]。また、前体と後体の境目は1枚の横隔膜(diaphragm)に分かれ、これも呼吸に関与する構造だと考えられる[17]。ヘモシアニンなどの呼吸色素をもたない[31]。
神経系
[編集]ヒヨケムシの前体の中枢神経系はクモガタ類として典型的で、眼・鋏角・触肢に対応する3節の脳神経節(食道上神経節 supraoesophageal ganglion とも、前大脳 protocerebrum・中大脳 deutocerebrum・後大脳 tritocerebrum を含む)と脚に対応する4節の神経節(食道下神経節 suboesophageal ganglion)が1つの集中部(synganglion)となっている[17][44]。第4脚のラケット器官に連結した特殊な神経があり、これは触肢神経の上方まで前へ伸びていた[44]。後体の中枢神経系は退化的で、1節の神経節のみをもつ[26][17]。
消化系
[編集]ヒヨケムシの消化系は発達で、順に前腸・中腸・後腸に分けられる[17]。前腸は口上板などに覆われる咽頭と、脳の間(食道孔 oesophageal foramen)を貫通する食道からなる[26][17]。中腸は脳の後方から後体の末端直前まで長大に伸ばし、前体と後体の境目でくびれている[26][17]。両筋に発達した消化腺(diverticula)が並び、特に後体のものは体腔を充満した数多くの分岐に枝分かれている[26][17]。肛門直前の後腸は、排泄物を一時的に貯まる用の嚢を背面にもつ[26][17]。
循環系
[編集]ヒヨケムシは節足動物として典型的な循環系(背面にある縦長い心臓)をもつが、それ以上に詳細な情報は限られている。19世紀の古典的な記載によれば、心臓は前体後半から後体後半まで伸ばし、前体に2対、後体に6対の心門(ostium)をもつとされる[17]。
排出器
[編集]ヒヨケムシの排出器官である基節腺(coxal gland)はコヨリムシ類と同様、触肢基節のみに開口し、途中にもう1つの嚢がある。これは他のクモガタ類における脚(通常第1脚基節のみ、一部の群は第1と第3脚基節)のみに開口し、末端の嚢のみをもつ基節腺とは明らかに異なる[53]。
前体の筋肉と内骨格
[編集]節足動物の頭部や前体は原則として内腹板(内腹甲、endosternite, 消化管と腹神経索の間に備わり、付属肢の基部を動かす筋肉に付着される軟骨性な内骨格)をもつが、ヒヨケムシは内腹板が退化消失し、代わりに第2脚基節に支えられる腱がアーチ状の内骨格に発達して消化管と腹神経索の間で会合し、脚の転節(ヒヨケムシにおいては最初の可動肢節)を動かす筋肉に付着される[30]。この点は他の多くのクモガタ類(例えばクモは腹板に支えられ、脚の可動な基節を動かす外在筋に付着される内腹板をもつ)とは随分と異なり、ヒヨケムシの転節を動かす筋肉が外在筋のように、基節の腱が内腹板のように機能すると言われている[30]。
毒の有無
[編集]確定的な記録に限れば、ヒヨケムシはクモやサソリのような注入性の毒(venom)はなく、自身を不味くするような防御物質(poison)ももたず[35]、全般的に無毒とされる[20][17]。
なお、インドのタミル・ナードゥ州に生息するヒヨケムシの1種 Rhagodima nigrocincta (=Rhagodes nigrocinctus) に関しては、Aruchami & Sundara Rajulu 1978 で鋏角両指が毒腺に特化した上皮腺があると報告されている。この論文によれば、その毒はセロトニン(一部のサソリやハチの毒にも見られる成分)を含んでおり、抽出してトカゲに注入したところ、10匹のうち7匹が麻痺したとされる。しかしこの種についての追試が行われておらず、他のヒヨケムシからも毒は見つからない[12]。いずれにせよ、この不確実な記録だけでヒヨケムシ全般を有毒生物というのは不適切である[12]。
生理学と生態
[編集]ヒヨケムシは研究が少ないため、詳細な生理学と生態は、ごく一部の種類のみによって知られている[20][12][54][13]。
生息環境と活動様式
[編集]
ヒヨケムシは主に砂漠やその周辺の荒れ地に生息し、そこの昆虫などの捕食者である同時に鳥類などの主な獲物でもあるため、乾燥帯の生態系における重要なニッチ(生態的地位)を占めると考えられる[55][56][17][12]。例外としてヒカラビヒヨケムシ科の構成種は植物の多い地域を好む傾向があり[54]、Dinorhax rostrumpsittacici は現世で唯一に熱帯雨林に生息するヒヨケムシである[4][29]。
多くの種類は夜行性で、名に現れるように日当たりを避けている。しかし日中で活動する昼行性種もあり、ウモウヒゲヒヨケムシ科、ヒカラビヒヨケムシ科とヒヨケムシ科で特に多く知られている[12][54][4]。ヒヨケムシ科の昼行性種からは、日中で後体を立てて体温調節をする行動が観察される[57]。また、雨など湿度の高い天候を嫌がるらしく、水をかけられるとすぐ穴を掘って身を隠す[45]。寿命が数年ほど長い種類は、一年中に夏を中心にして外で活動し、残りの時期を巣穴に隠れて休眠する[54]。
活動的で、徘徊する間では獲物や配偶に遭うまで立ち止まらないほどである[7]。「風のように走る」と言われるほど素早く走り、最速のものはおよそ時速16kmまで達する[58]。特に体毛が発達したものは、走る姿が転んでいる冠毛にも見えて[5]、ネズミと見間違われることすらある[20]。触肢の吸盤を利して滑らかな表面を掴み、植物やガラスなどもある程度登れるが、基本的には地表で活動する[45]。移動の際、第2-4脚のみ歩脚として歩行に用いられ、第1脚と触肢は地面から浮かんで感覚器として働いている[40]。第4脚のラケット器官の明確な機能は未だに不明だが、餌や配偶を探すのに役立つ化学受容器とされ、またサソリの感覚器である櫛状板のように常に地面に触れるため、それに似た機能(振動を読み取るなど)をもつとも考えられる[59][44]。
その高い活動性を支えるように、ヒヨケムシの代謝率は高く、発達した気管系で効率よく呼吸する。呼吸色素をもたないため、ガス交換は完全に気管系に依存すると考えられる[31]。昆虫のように気門を開閉して断続的な換気を行い[54]、前体の後半が常に上下の伸縮を繰り返すのは、横隔膜の伸縮による換気の動きだと考えられる[17]。大型種の鋏角に見られる気嚢は昆虫のそれに似て、酸素を蓄える、もしくはふいごのように伸縮して能動的な換気を行ったと考えられる[31]。ヒヨケムシの発達した気管系は、前述のようなガス交換の効率を上げるだけでなく、エネルギーの節約に繋がる軽量化にも役立つと考えられる[31]。特に大型種が鋏角に気嚢をもつのは、この強大な捕食器官の酸素供給と軽量化に適した性質だと考えられる[31]。
走光性
[編集]夜行性種は昼では日当たりを避けているが、一部では夜中の人造光源に引き寄せられ、正の走光性があるように見える。しかしこれは正の走光性をもつ夜行性昆虫のように、元々自然光源(天体の光)でナビゲートする習性をもち、人造光源をそれと勘違いしたからではなく、人造光源に集まった獲物である昆虫に引き寄せられた、もしくは明暗のコントラストを獲物の出没と勘違いていたからだと考えられる[60]。
巣穴
[編集].jpg)
ヒヨケムシは全般的に自力で巣穴を掘ることができる[45]。巣穴は円形から楕円形で、外で活動しない時間(摂食・脱皮・産卵・夜行性種の日中・昼行性種の夜)をそこで過ごす[45][57][54]。材質は土壌から植物組織まで、人工物まで含むと木材や厩肥も知られている[45]。土で作られた巣穴は地下、もしくは小石・丸太・牛糞など大きめな固形物に覆われている[45]。巣穴を作る際には、鋏角で固まった土を粉砕し、第2-3脚脛節の棘で緩い土を体の腹面から後方に飛ばし、鋏角・触肢・第1脚で掘り出した土を入り口の外まで押し出す[45]。また、土の代わりに木(朽ち木など)に巣穴を作る[45]・巣穴の構造が雌雄により異なる[12]・他の動物から脱落した毛を巣穴の裏地にする[6]、などの習性をもつ種類も知られている。
餌と天敵
[編集]

ヒヨケムシは俊敏に地表を徘徊する活動的な捕食者であり、他の節足動物(昆虫と他のクモガタ類)を主食とし、小型脊椎動物(ネズミ・トカゲ・ヘビ・鳥類など)を捕える場合もある[20][61][62]。種類・雌雄・成長段階によりある程度の好みの違いが見られるが、原則として偏食性をもたず、制圧できる大きさの小動物であれば捕食するジェネラリストである[63][45][61][62]。様々な獲物の中で、防御物質(毒など)をもつもの(アリ・アリバチ・ヤスデなど)が最も一般に拒否され、特にヤスデは捕食されても無毒な部分(頭部と前4胴節)だけ食られ、有毒な部分(胴部のほとんど)が捨てられる[62]。一部の種類はアリを自発的に襲いかかる行動が観察されるが、食べずに殺しているため、アリ食とは別の目的による行動の可能性がある[7]。代謝率は高いため、頻繁に餌を摂る必要があるとされる[7]が、1ヶ月以上の絶食に耐える種類も知られている[45]。水を嫌がるため、普段の獲物から充分な水分を摂取できていると考えられる[45]。また、自然な状況ではないが、飼育下の観察で昆虫の遺骸や人工的に用意された餌(加水分解酵母・牛ひき肉など)を受動的に摂食するとこと[63]と、ロードキルされた爬虫類の遺骸を摂食することも観察される[64]。

獲物を探す際には、周りを感知するように徘徊しながら触肢を振りまわして、触覚・視覚・振動に頼って獲物を発見していたと考えられる[63][45][65]。触肢先端の吸盤を出して獲物を掴み[37]、強力な鋏角でそれを補殺して咀嚼する。その過程で後体は伸縮し、鋏角はすり合わせるように左右相互にはさみを開閉しながら前後に動く[63]。獲物の体液を鋏角直後の口で吸収しながら、口上板前端のブラシ状の剛毛で不要な物質を濾過したと考えられる[20][32][17]。また、成体は自発的に徘徊しながら獲物を探すのに対して幼生は待ち伏せして獲物を捕るという、同一種類が成長段階の違いによって異なった捕食行動をとるものもある[66]。
.jpg)
ヒヨケムシの主要な天敵は脊椎動物であり、同じ生息地の哺乳類・爬虫類・鳥類などの糞からその鋏角の残骸が多く見られ[67][62]、主食ほど大きな割合(約40%)を占める例もある[12]。他の節足動物捕食者としてクモ・サソリ・別種のヒヨケムシなどが知られるが、ヒヨケムシを主食にした、それとも単に偶発的なのかは判断しにくい[67]。明らかにヒヨケムシを狙って捕食する例は、ナミビアに生息し、地面にトンネル状の巣穴を作るアシダカグモ類の一種が挙げられる。このクモは何らかのメカニズムでヒヨケムシ科の1種 Metasolpuga picta の雄を自らの巣穴に同種の雌が居ると勘違いさせて、巣穴まで誘惑して捕食することが知られている[57]。
飼育下では配偶行動などがもたらす共食いが高頻度に観察される[63][57][12][68]が、自然環境では交接後の雄が無事に逃げ出すことができ、配偶行動以外でも同種が触れ合うと積極的にお互いを避けるため、実際の共食いの頻度は低かったと考えられる[63][57]。
防御と擬態
[編集]

危険に遭うと触肢を高く上げ、鋏角を開き、後体を立てて威嚇する場合がある。それでも相手が諦めない場合は、鋏角で噛み付いて自衛する。背面に向けて発達した中眼は敏感な視力を有し、これは天敵への警戒に役立つと思われる[57]。また、鋏角を擦り合わせ、摩擦音を出して威嚇する種もある[69]。
ナミビアに生息するヒヨケムシ科の一種は、黒い体に白い背板をもつ。これは黒い体と白い前翅をもつ一部の Onymacris 属と Stenocara 属のゴミムシダマシ科の甲虫に防御擬態する特徴だと考えられる[70]。
ヒヨケムシが別生物の擬態モデルとなった例もある。例えばイランに生息するヘビの1種 Pseudocerastes urarachnoides は、尻尾の先端がヒヨケムシの後体と脚に攻撃擬態した構造で、それを用いて鳥類を誘惑して捕食する[70]。
繁殖と発育
[編集]
.jpg)
ヒヨケムシの精子のやり取りは多くのクモガタ類と同様、真の交尾ではなく、精包の受け渡しを通じて行う「交接」となっている。なお、ヒヨケムシは唯一に鋏角で精子の受け渡しをする鋏角類であり[4]、交接中の雌が硬直する・雄が雌に噛み付くなど、独特な配偶行動もいくつか知られている[45][13]。しかし2021年現在、12科約140属約1,100種のうち5科9属17種[注釈 13]のみ詳細な繁殖行動が記載されるため、全面的な情報は限られている[13]。
配偶行動
[編集]ヒヨケムシの雌雄のうち、雄が自発的に配偶を探す側である。少なくともヒヨケムシ科の場合、雄は地中の巣穴に隠れた雌の位置も正確に判断できるため、フェロモンで辿った可能性がある[57]。雄が雌を見つかると、鋏角や触肢で雌に接触しようとする。多くの場合、雌がこの際に雄を受け入れれば麻痺のように硬直状態(frozen state, torpor, quiescence)に入り、生殖孔を露出させるように後体を反り上げる[71]。雌のこの反応を誘発する要因は不明だが、雄の接触によると考えられ、人為的な刺激を受けてもこの状態に入ることがある[72][13]。また、硬直の程度は種類によって異なり、例えばサメヒヨケムシ科とヒヨケムシ科は完全硬直、ヒトリヒヨケムシ科は種類により完全から部分的硬直(付属肢が僅かに動く)、スナハシリヒヨケムシ科とコヒヨケムシ科は硬直しない[68][13]。また、交接直前の雄は、種類により鋏角で雌の体(多くは生殖孔、種類により後体側面や脚の基部まで)をマッサージする[73][68]・触肢で雌の背面を小刻みに叩く・硬直中の雌を持ち上げながら歩く[74]などの行動も観察され、特に雌が硬直しない種類は、雄が触肢や脚で雌の動きを制限する[68][13]。雄の触肢と脚の性的二形な長さは、これらの行動で役に立つ(雌をより上手く掴める・凶暴な雌を一定の距離を保ちながら触れるなど)[13]。
交接の際、雄は事前に生殖孔から排出した精包を鋏角で雌の生殖孔に押し込む[73]。多くの場合、雄は排出した精包を何らの表面(多くの場合は地面、ヒヨケムシ科は雌の背面)に置いてから鋏角で拾い上げるが、ヒトリヒヨケムシ科の場合、雄は生殖孔から精包を直接的に雌の生殖孔に置いてから鋏角で押し込む[9][73][71][13]。
雄の鋏角で雌に触れる上述の行動は、多くが咀嚼のように乱暴で、雌に傷つける危険性を伴っている[63][68]。多くの種類において雄の鋏角が相対的に貧弱であったのは、これらの配偶行動で雌を傷つけないように適応した特徴だと考えられる[57]。
雄の鞭毛複合体は交接の際に雌の生殖孔内部まで入れるため、交接において何らかの重要な役割を果たした器官と考えられる。しかしその正確な機能は未だに不明で、物を把握できるような特殊な構造により移精器官(精子を雌の生殖腺内まで押し込むのに役立つ)説が特に有力視されるが、他の雄の精子を除去/破壊する・雌に何らかの必要な刺激を与えるなどの説もある[75]。
精包が完全に雌の生殖孔に収まれる度に交接は終了し、硬直状態の雌はここで徐々に再活性化し、雄はこの隙に素早く去っていく[13]。ヒヨケムシ科の場合、交接後の雄は鋏角を砂に入れて綺麗にするような行動が観察される[57]。雄は複数回の交接を行えるが、雌はそうとは限らない(例えばコヒヨケムシ科は1回のみ)[13]。なお、雄の精子は有限(成体の精巣は萎縮し、精管に蓄えた精子はそれ以前に形成されたもの)で、交接後の寿命も短い(1週間から1ヶ月)ため、長くは続かないと考えられる[13]。また、精子は無鞭毛で自力で動けないため、何らかの外力(雄の鋏角の動き・雌の生殖腺の蠕動などと推測される)に頼って卵子にたどり着いて受精させたと考えられる[13]。
雄が交接を拒否する雌に捕食される・交接中の雌が雄に傷つけられて捕食される・交接後の雄が逃走失敗して再活性化/制圧から解放された雌に捕食される、などの性的共食いが知られるが、どれも飼育下による記録のため、実際の生態を反映しない可能性がある[13]。例えばヒヨケムシ科の1種 Metasolpuga picta の場合、自然環境では雄がどれも無事に逃げ出し、このような共食いが観察されなかった[57]。
生活環
[編集]産卵の際、雌は深い巣穴を掘って丸い卵を産む。卵の数は種類により30から200個の間である[54]。育児習性をもつ雌の場合、卵を保護する間に捕食はしない為、産卵の前に大量の餌を摂る[76][77]。サメヒヨケムシ科の場合、産卵から孵化までの胚発生は多くが1-2週かかるが、1-2日だけで孵化する種類もある[13]。生まれたての幼生(post-embryo)は歩けなくて単調な姿をしており、1回目の脱皮を経て歩ける成体の姿に近い1齢幼生となる。少なくとも一部の種類は、1齢幼生まで親の巣穴で群れに集まって過ごし、2齢幼生になってから巣立ちをする[77]。ほとんどの種類の齢期数は不明だが、知られる限り(post-embryo 期を除いて)4から10齢期に及ぶ[54]。成体以降は脱皮せず[54]、雄の鋏角鞭毛複合体は成体になってから現れる[4][78]。寿命は種類により1年から5年以上[54]、雄の方が短命と考えられる[20][77]。生活環は種類により1年から数年周期である[54]。
分布と分類
[編集]ヒヨケムシは熱帯と亜熱帯に分布する。現生種はほぼ全てが砂漠やその周辺の乾燥地帯に生息し、熱帯雨林に生息するのは1種(Dinorhax rostrumpsittacici)のみ知られている[29]。なお、オセアニアとマダガスカルの乾燥地帯には分布しない[9]。また、クモガタ類にしては珍しく、洞窟性の種類は知られていない[9]。
何らかの理由で逃亡した個体が発見された例があるが、日本には分布しない[79]。
系統位置
[編集]

鋏角類の基本体制が未解明な19世紀頃では、ヒヨケムシはその一見して昆虫的な性質(頭部と胸部に似た前体・発達した気管系など)により、「クモガタ類と昆虫の中間型」などと誇張に解釈されることもあった[25]。クモガタ類を含む鋏角類は昆虫とは別系統だと判明した20世紀以降では、ヒヨケムシは疑いなく真鋏角類のクモガタ類に含める鋏角類であり、その単系統性も形態学・分子系統学の両方の系統解析に立証される[80][81][82][83][84]。しかし多くのクモガタ類の目と同様、ヒヨケムシの真鋏角類における系統位置は2010年代以降では不明確である[85]。
20世紀から2000年代まででは形態学上の多くの共通点、例えば背甲と関節した2節のはさみ型の鋏角・短い膝節らしき肢節をもたない(短い腿節と長大な膝節をもつ)脚・後体第3-4節にある気門などに基づいて、ヒヨケムシは一般にカニムシに最も近縁で姉妹群(共に Haplocnemata/Apatella をなす[86])と考えられた[87][33][81]。しかし2010年代以降、これは多くの分子系統解析に否定され、代わりにクツコムシ類の姉妹群[88][89]・胸板ダニ類(Acariformes)の姉妹群[90][91](共に Poecilophysidea をなす[27][86])・胸板ダニ類やコヨリムシ類と単系統群になる[90](共に Cephalosomata をなす[27][86])・カブトガニ類+クツコムシ類+蛛肺類の姉妹群[92]・カブトガニ類+クツコムシ類の姉妹群・ダニ類の姉妹群[93]・コヨリムシ類の姉妹群[53]など、様々な不確かな解析結果が出ている。
もしヒヨケムシは本当に胸板ダニ類やコヨリムシに近縁であれば、ヒヨケムシにおけるいくつかの性質(これらの群に似た前体の分節、もしくはコヨリムシに似た基節腺の構造)は、この類縁関係を表した共有派生形質かもしれない[53]。また、ヒヨケムシの雄性生殖腺の細部と精子の構造も、カニムシより胸板ダニ類に似ている[51][43]。
下位分類
[編集]ヒヨケムシの中で、現生は12科、十数亜科、約140属と1,100以上の種[4]、確定的な化石は単形の5属5種が記載される(そのうち2種科未定、後述参照)[94]。ほとんどの科は旧世界(アジア・アフリカ・ヨーロッパ)と新世界(アメリカ大陸)のいずれかのみに分布する[9]。クモガタ綱の目のなかで、ヒヨケムシの種数は6番目に多い[85][95]。

数多くの科・亜科・属・種は20世紀中期で Carl Friedrich Roewer によって記載と創設をなされ、ヒヨケムシの下位分類体系の基盤を立ち上がった[26]。しかしその分類体系の指標とされた一部の性質、特に歩脚跗節の棘は欠損しやすくて同種や同個体ですら違いが見られる(すなわち類縁関係を反映しない可能性が高い)にもかかわらず、一部の科と多くの属の主要な指標とされていた[12][4]。Roewer のこのような分類体系は「紛らわしい」や「混沌」と批判されほど懐疑的で、その後の研究者を悩ませる難題となっている[12][4]。また、鋏角の特徴は昔今を通じて種の同定形質として重要視されるにもかかわらず、各部位(特に雄の鞭毛複合体)の用語と定義が文献記載によって異なり、混乱を招きやすく、相同性が検討しにくい[4]。
2010年代では、暫定的に次の12科の構成と分布域が認められる(特記しない限り和名は小野 2002[16]、化石種は Dunlop et al. 2020[94]、その他の情報は Bird et al. 2015[4] に基づく)。
| 学名 | 和名 | 亜科 | 現生属 | 現生種 | 化石種 | 分布域 |
|---|---|---|---|---|---|---|
| Ammotrechidae | スナハシリヒヨケムシ科 | 4 | 24 | 104[96] | 1 | 新世界:アメリカ南部、メキシコ、カリブ海地域、南アメリカ |
| Ceromidae | ミナミヒヨケムシ科 | 3 | 20 | 1 | 旧世界:南部アフリカ、東部アフリカ 新世界:ブラジル | |
| Daesiidae | コヒヨケムシ科 | 6 | 29 | 190 | 1 | 旧世界:アフリカ、ヨーロッパ南部、中東 新世界:アルゼンチン、チリ |
| Eremobatidae | ヒトリヒヨケムシ科 | 2 | 8 | 187 | 新世界:カナダ南部、アメリカ西部(ミズーリ川 - ミシシッピ川周辺)、メキシコ | |
| Galeodidae | サメヒヨケムシ科 | 9 | 200[97] | 旧世界:北部アフリカ、中東、中央アジア、インド亜大陸 | ||
| Gylippidae | アジアヒヨケムシ科 | 2 | 5 | 27 | 旧世界:近東、南アフリカ | |
| Hexisopodidae | アナホリヒヨケムシ科 | 2 | 23 | 旧世界:南アフリカ | ||
| Karschiidae | カルシュヒヨケムシ科 | 4 | 45 | 旧世界:北アフリカ、中東、中央アジア | ||
| Melanoblossiidae | ウモウヒゲヒヨケムシ科 | 2 | 6 | 17 | 旧世界:南アフリカ、東南アジア(インドネシア、ベトナム) | |
| Mummuciidae | ヒカラビヒヨケムシ科 | 8 | 28[98] | 新世界:南アメリカ | ||
| Rhagodidae | オオヒヨケムシ科 | 27 | 98 | 旧世界:北アフリカ、中東、アジア、インド亜大陸 | ||
| Solpugidae | ヒヨケムシ科 | 2 | 17 | 191 | 旧世界:アフリカ、中東 |
前述のような問題から大きな影響を受けて、ヒヨケムシの内部系統関係は研究しにくく、2010年代以降でも再検討が必要とされる科や属が多く残される[4]。Roewer の記載では第2-4脚跗節の節数を基に、跗節が分節しないオオヒヨケムシ科・アジアヒヨケムシ科・カルシュヒヨケムシ科は基盤的で、4節以上に分かれた跗節をもつスナハシリヒヨケムシ科・コヒヨケムシ科・ヒヨケムシ科は最も派生的だと考えられた[26]。なお、2010年代の分子系統解析では様々な相容れない解析結果が出て、特にスナハシリヒヨケムシ科とアジアヒヨケムシ科は多系統群とされる場合もある[85][95]。Kulkarni et al. 2023 の解析によると、ヒヨケムシの内部系統はローラシア大陸産のBoreosolifugae[注釈 14]と、ゴンドワナ大陸産のAustralosolifugae[注釈 15]という2つの亜目で大きく分かれている[99]。
2000年代以降では、雄の鋏角鞭毛複合体の基本形態・第1脚の爪の数・第2-4脚跗節の節数・背甲両前端の外骨格の癒合具合・触肢腹面の棘・気門周辺の突起物・肛門の位置が科と亜科を区別するのに有用な同定形質とされる[9][4]。種は主に鋏角の歯・雄の鋏角鞭毛複合体などの細部形態を基に区別される[4]。なお、サメヒヨケムシ科とオオヒヨケムシ科のように、全般的に体色の方が種特異性が顕著なものもある[4]。
スナハシリヒヨケムシ科 Ammotrechidae
[編集]
a: Neocleobis solitarius, b-c: Ammotrechona 属, d-f: Ammotrecha 属
-
 Ammotrecha sp.
Ammotrecha sp. -

-

-

.jpg)



| |||||||||||||||||||||
| Iuri et al. 2021 に基づいたスナハシリヒヨケムシ科の内部系統関係[100] |
現生4亜科24属104種(Iuri & Iglesias 2022 時点)[96]、化石1属1種(Dunlop et al. 2020 時点)が記載される[94]。
新世界のアメリカ南部、メキシコ、カリブ海地域と南アメリカから発見される[4]。
雄の鋏角鞭毛は不動、膜状、楕円形の椀状[4]。第1脚跗節に爪は無く、第2-4脚跗節の節数は1・1・1から2・2・4[42]。



現生3属20種(Bird et al. 2015 時点)[4]、化石1属1種(Dunlop et al. 2020 時点)が記載される[94]。
現生種は旧世界の南部アフリカと東部アフリカ、化石種は新世界のブラジル(セアラー州)から発見される[4]。
(鞭毛をもたない Toreus 属を除いて)雄の鋏角鞭毛は可動、上に曲がり返した鞭状、基部は3-8本の集約した剛毛に覆われている[4]。第1脚跗節は爪2本、第2-4脚跗節の節数は2・2・2[42]。
| 属 |
|---|
| Ceroma |
| Ceromella |
| Cratosolpuga† |
| Toreus |


-

-

-
 コヒヨケムシ科の1種
コヒヨケムシ科の1種 -





現生6亜科29属190種(Bird et al. 2015 時点)[4]、化石1属1種(Dunlop et al. 2020 時点)が記載される[94]。
旧世界のアフリカ、ヨーロッパ南部と中東、および新世界の南アメリカ(アルゼンチンとチリ)から発見される(唯一に現生種が旧世界と新世界の両方に分布する科)[4]。
雄の鋏角鞭毛は可動、上に曲がり返した膜状から鞭状[4]。第1脚跗節に爪は無く、第2-4脚跗節の節数は1・1・1から2・2・4[42]。
元々は Kraepelin 1899 によってヒヨケムシ科の1亜科 Daesiinae として創設されたが、Roewer 1934 以降では独立の科として区別されるようになった[26][4]。
Muma 1971 で記載された Amacata penai は独自にチリヒヨケムシ科 Amacataidae に分類された[102]が、Maury 1985 によりコヒヨケムシ科の1種 Ammotrechelis goetschi のシノニムとされるに連れて、チリヒヨケムシ科もコヒヨケムシ科のシノニムとされるようになっている[103]。
ヒトリヒヨケムシ科 Eremobatidae
[編集]
-
 ヒトリヒヨケムシ科の1種の雄
ヒトリヒヨケムシ科の1種の雄 -
 ヒトリヒヨケムシ科の1種の雄
ヒトリヒヨケムシ科の1種の雄 -
 ヒトリヒヨケムシ科の1種
ヒトリヒヨケムシ科の1種 -
 ヒトリヒヨケムシ科の1種
ヒトリヒヨケムシ科の1種
.jpg)
_(28659789713).jpg)
.jpg)
.jpg)
| |||||||||||||||||||||||||||||||||||||||
| Cushing et al. 2016 に基づいたヒトリヒヨケムシ科の内部系統関係[85] |
現生2亜科8属187種が記載される(Bird et al. 2015 時点)[4]。化石記録なし(Dunlop et al. 2020 時点)[94]。
新世界のカナダ南部、アメリカ西部(ミズーリ川 - ミシシッピ川周辺)とメキシコから発見される[4]。
雄の鋏角鞭毛複合体は集約した数本の剛毛、鞭毛は可動、前向きの針状(鞭毛をもたない種類もある)[4]。第1脚跗節は爪1-2本、第2-4脚跗節の節数は1・1・1から1・1・3[42]。
雄の鋏角不動指が細く、往々にして滑らかな針状に特化している[104][4]。精包は直接に雄の生殖孔から雌の生殖孔に付けられる[72][71]。
Chanbria・Horribates・Eremorhax 以外の属、および2亜科とも非単系統群だと示される[85]。
サメヒヨケムシ科 Galeodidae
[編集]



).jpg)
現生9属200種が記載される(Erdek 2021 時点、種数最多の科)[97]。化石記録なし(Dunlop et al. 2020 時点)[94]。
旧世界の北部アフリカ、中東、中央アジアとインド亜大陸から発見される[4]。
雄の鋏角鞭毛は後向きの匙状[4]。第2-4脚跗節の節数は2・2・3[42]。
爪の基部節に細かな剛毛が並び、気門を守るような櫛状の構造体がある[4]。大型種が多く、最大級のヒヨケムシが含まれる[106]。ほとんどの種は Galeodes 属に含まれる[4]。Paragaleodes 属は全てが昼行性である[4]。
| 属 | 種数 |
|---|---|
| Galeodes | 174[97] |
| Galeodopsis | 5 |
| Galeodumus | 1 |
| Gluviema | 1 |
| Othoes | 4 |
| Paragaleodes | 12 |
| Paragaleodiscus | 1 |
| Roeweriscus | 1 |
| Zombis | 1 |
アジアヒヨケムシ科 Gylippidae
[編集]

).jpg)

現生2亜科5属27種が記載される(Bird et al. 2015 時点)[4]。化石記録なし(Dunlop et al. 2020 時点)[94]。
雄の鋏角鞭毛複合体は亜科によって異なる。Gylippinae 亜科は鞭毛が不動、前向きのひも状、基部から途中まで不動指背面と癒合し、掌部側背面に1本の強大な剛毛(retrolateral manus spiniform seta)をもつ。Lipophaginae 亜科は前向きに集約した数本の剛毛(鞭毛なし)[4]。第2-4脚跗節の節数は1・1・1[42]。
| 属 | 所属亜科 | 種数 |
|---|---|---|
| Acanthogylippus | Gylippinae | 1 |
| Gylippus | Gylippinae | 19 |
| Lipophaga | Lipophaginae | 3 |
| Trichotoma | Lipophaginae | 3 |
| Bdellophaga | Lipophaginae | 1 |
アナホリヒヨケムシ科 Hexisopodidae
[編集]

-
 Chelypus sp.
Chelypus sp. -

-

-


).jpg)
).jpg)

現生2属23種が記載される(Bird et al. 2015 時点)[4]。化石記録なし(Dunlop et al. 2020 時点)[94]。
雄の鋏角鞭毛は可動、螺旋状に曲がり返した鞭状、基部はカップ状で不動指基部近くに付く[4]。第2-4脚跗節の節数は1・1・1[42]。第4脚は第3脚より短く、跗節に爪は無い[26][42][4]。ラケット器官は2-5対[26]。
地下性に特化したずんぐりな体型が特徴的。脚は短縮して丈夫な棘が並び、胴体は丸くて分厚い剛毛に覆われている。その特徴的な外見から「mole solifuges」("モグラヒヨケムシ")や「teddybear solifuges」("テディベアヒヨケムシ") などと呼ばれている[43][108]。また、地下性のため見つかりにくく、実際の種数はもっと多かったと考えられる[4]。
| 属 | 種数 |
|---|---|
| Chelypus (=Siloanea) | 8 |
| Hexisopus (=Mossamedessa) | 15 |
カルシュヒヨケムシ科 Karschiidae
[編集]


現生4属45種が記載される(Bird et al. 2015 時点)[4]。化石記録なし(Dunlop et al. 2020 時点)[94]。
雄の鋏角鞭毛は不動、螺旋状に曲がり返した鞭状もしくは2本の扇形膜状[4]。第2-4脚跗節の節数は1・1・1[42]。
鋏角不動指の歯は原則として同規的で数多く(multidentate)、摩擦音を出す用の構造体をもたない[4]。気門は櫛状に並んだ剛毛に覆われている[4]。一部の種類は雄の鋏角不動指背面に1本の突起が生えて、不動指全体が二股状となっている[4]。
| 属 | 種数 |
|---|---|
| Barrus | 2 |
| Barrussus | 2 |
| Eusimonia | 15 |
| Karschia | 26 |
ウモウヒゲヒヨケムシ科 Melanoblossiidae
[編集]

現生2亜科6属17種が記載される(Bird & Wharton 2015 時点)[4][109]。確定的な化石記録なし[94](Dunlop et al. 2020 時点。ただし科未定化石種 Cushingia ellenbergeri は本科の Dinorhax 属に近い性質をもつ[110])。
旧世界の南アフリカと東南アジア(ベトナム・インドネシア)から発見される[4]。
雄の鋏角鞭毛複合体は亜科によって異なる。Dinorhaxinae 亜科は鞭毛が不動、下向きの鞭状、基部が不動指先端付近に付く[4][109]。Melanoblossiinae 亜科は不動指の基部内側に集約した数本の剛毛、鞭毛は前向きの羽毛状(鞭毛をもたない種類もある)[4][109]。第2-4脚跗節の節数は1・1・1から1・1・2[42]。
他の性質も亜科によって大きく異なる。Dinorhaxinae 亜科は東南アジア産で比較的大型(4cm程度)、全体的にオオヒヨケムシ科によく似ている(肥厚な鋏角・丈夫な脚・腹面に開く肛門などをもつ)のに対して、Melanoblossiinae 亜科は南アフリカ産で小型(1cm以下[注釈 19])、鋏角は細い[4]。なお、両亜科とも昼行性とされる[4][29]。
本科における Dinorhaxinae 亜科の唯一の種かつ既知唯一の東南アジア産現生ヒヨケムシでもある Dinorhax rostrumpsittacici は、最初では Simon 1877 でオオヒヨケムシ科 Rhax 属の種 Rhax rostrumpsittaci として記載された。本種は Simon 1879a 以降では新属 Dinorhax の種 Dinorhax rostrumpsittacici とされ、Roewer 1923 により本科の新亜科 Dinorhaxinae に再分類された[26]。しかし、本亜科は前述の通り Melanoblossiinae 亜科よりオオヒヨケムシ科に似るため、それを含んだ本科は多系統群である可能性が高い[4][29]。
ヒカラビヒヨケムシ科 Mummuciidae
[編集]
b: Gaucha stoeckeli
c: Metarleobis fulvipes
d: Mummucipes paraguayensis

現生8属28種が記載される(Souza et al. 2021 時点)[98]。化石記録なし(Dunlop et al. 2020 時点)[94]。
雄の鋏角鞭毛は膜状、袋状で前端に開口をもつ[4]。第2-4脚跗節の節数は1・1・1から1・1・2[42]。
前体後半から後体全長にかける背面の縦縞模様(背板中央と体節両背面黒/暗褐色、背板両縁白)が特徴的[22]。昼行性で、砂漠や荒れ地より植物の多い地域を好んで生息する[54]。小型種に構成され、知られる中で最小(3.9 - 5.85mm)のヒヨケムシ Vempironiella aguilari を含む[22][注釈 19][4]。
元々は Roewer 1934 によってスナハシリヒヨケムシ科の1亜科 Mummuciinae として創設されたが、Maury 1984 以降では独立の科として区別されるようになった[4]。
| 属 | 種数 |
|---|---|
| Cordobulgida | 1 |
| Curanahuel [111] | 1 |
| Gaucha (=Gauchella, Metacleobis)[112] | 13[98] |
| Mummucia | 5 |
| Mummucina | 5[113] |
| Mummucipes | 1 |
| Uspallata | 1 |
| Vempironiella [22] | 1 |
オオヒヨケムシ科 Rhagodidae
[編集]

-
 オオヒヨケムシ科の1種
オオヒヨケムシ科の1種 -
 オオヒヨケムシ科の1種
オオヒヨケムシ科の1種 -

-




現生27属98種が記載される(Bird et al. 2015 時点)[4]。化石記録なし(Dunlop et al. 2020 時点)[94]。
旧世界の北アフリカ、中東、アジアとインド亜大陸から発見される[4]。
雄の鋏角鞭毛は不動、前上方に湾曲した上下2本の短い鞭毛が円筒状の複合体(diploflagellum)をなす[4]。第1脚跗節は爪2本、第2-4脚跗節の節数は1・1・1[42]。
重厚な体型が特徴的。脚は丈夫で短く、鋏角は肥厚で雌雄とも歯が発達[4][114]。後体末端は大きな半円形で肛門は腹面に開く[42]。派手な体色をもつ種類も多い[114]。
ほとんどの属は未だに Roewer の懐疑的な体系に基づいままのため、全面的な再検討が必要とされる[4]。
| 属 | 種数 |
|---|---|
| Rhagodalma | 1 |
| Rhagodax | 1 |
| Rhagodeca | 3 |
| Rhagodelbus | 1 |
| Rhagoderma | 3 |
| Rhagoderus | 1 |
| Rhagodes | 27 |
| Rhagodessa | 5 |
| Rhagodeya | 2 |
| Rhagodia | 4 |
| Rhagodima | 2 |
| Rhagodinus | 2 |
| Rhagodippa | 1 |
| Rhagodira | 3 |
| Rhagodista | 1 |
| Rhagoditta | 6 |
| Rhagodixa | 3 |
| Rhagodoca | 17 |
| Rhagodolus | 1 |
| Rhagodomma | 1 |
| Rhagodopa | 4 |
| Rhagodorimus | 1 |
| Rhagodorta | 1 |
| Rhagodospsus | 1 |
| Rhagoduja | 1 |
| Rhagodula | 1 |
| Rhagoduna | 4 |
ヒヨケムシ科 Solpugidae
[編集]
-

-

-
 ヒヨケムシ科の1種の雄
ヒヨケムシ科の1種の雄 -
 Ferrandia ferrandii の第2(a)- 3(b, c)脚脛節・蹠節・跗節、および第4脚跗節(d)
Ferrandia ferrandii の第2(a)- 3(b, c)脚脛節・蹠節・跗節、および第4脚跗節(d)

_1600.jpg)

現生2亜科17属191種が記載される(Bird et al. 2015 時点)[4]。化石記録なし(Dunlop et al. 2020 時点)[94]。
雄の鋏角鞭毛は不動、上に曲がり返した鞭状、基部は嚢状に膨らんで鋏角不動指背面と癒合する[4]。第1脚跗節に爪は無く、第2-4脚跗節の節数は4・4・6(Ferrandiinae 亜科)もしくは4・4・7(Solpuginae 亜科、ヒヨケムシの中で最多)[42]。
ほとんどの属は未だに Roewer の懐疑的な体系に基づいままのため、全面的な再検討が必要とされる[4]。
| 属 | 所属亜科 | 種数 |
|---|---|---|
| Ferrandia | Ferrandiinae | 1 |
| Metasolpuga | Solpuginae | 1 |
| Oparba | Solpuginae | 4 |
| Oparbella | Solpuginae | 7 |
| Prosolpuga | Solpuginae | 1 |
| Solpuga | Solpuginae | 34 |
| Solpugassa | Solpuginae | 6 |
| Solpugeira | Solpuginae | 2 |
| Solpugella | Solpuginae | 5 |
| Solpugema | Solpuginae | 29 |
| Solpugiba | Solpuginae | 4 |
| Solpugista | Solpuginae | 4 |
| Solpugisticella | Solpuginae | 1 |
| Solpuguna | Solpuginae | 5 |
| Solpugyla | Solpuginae | 10 |
| Zeria | Solpuginae | 59 |
| Zeriassa | Solpuginae | 17 |
化石種
[編集]
ヒヨケムシは化石に保存されにくく、化石種の発見例は非常に稀である[115][11]。一部の種類(Protosolpuga、Cushingia など)はほとんどが乾燥地帯に生息する現生種とは異なり、森林など湿度の高い環境に生息したと考えられる[115]。2020年まででは、次の確定的な5属5種、不確かな1属1種と1つの未命名標本が知られている[116][94]。
- Cushingia ellenbergeri
- Palaeoblossia groehni
- 新生代古第三紀始新世(約4,900万 - 4,400万年前[1])に当たる琥珀 Baltic amber(旧世界、ロシア、カリーニングラード州)から発見される[119]。コヒヨケムシ科に分類される[94]。
- Solifugae gen. et sp. indet.
- 新生代古第三紀始新世(約4,900万 - 4,400万年前[1])に当たる琥珀 Baltic amber(旧世界、ロシア、カリーニングラード州)から発見される。科・属・種未定とされる[116]。
- Happlodontus proteus
- 新生代新第三紀中新世(約1,600万年前[1])に当たる琥珀 Dominican amber(新世界、ドミニカ共和国)から発見される[101]。スナハシリヒヨケムシ科に分類される[94]。
人間との係わり
[編集]
ヒヨケムシは人間の日常とはさほど係わりのない動物である。しかし原産地では一部の夜行性種が人工光源に引き寄せられ、住宅に入り込むことがある[54]。稀にペットとして流通するが、素早い動きと夜行性な生態により、発見・採集・飼育・研究とも困難であり[120][60][71][49]、ヒヨケムシを中心に研究をする存命の学者も少ない[20]。なお、光源に引き寄せされる性質を利用し、ライトトラップとピットフォールトラップをあわせて採集することは可能である[120][60]。
過剰に刺激されると、自衛のため鋏角で噛み付くことがあり、これは強大なものであれば痛みを生じて傷つける場合もある[20][121]。その傷口を適切にケアしないと何らかの二次的症状が起きる場合があるが、これはヒヨケムシ自体ではなく、傷口に侵入した外界の病原体(細菌やウイルス)が原因である[20][121]。いずれにせよ、ヒヨケムシ自体は無毒で人間を自発的に襲うことはなく、無駄に刺激しなければ無害で、過剰に怖がる必要はない[18][19]。
誤解
[編集]
ヒヨケムシは基本的に無害な動物であるにもかかわらず、その凶暴な見た目や様々な言伝え・逸話・噂・迷信・誇張・都市伝説・デマなどにより、過剰に曲解・畏怖されることが多い[18][19][8][121]。例えばイランでは、ヒヨケムシは人を殺せるほど危険で、常に人を襲うとする邪悪な生物という迷信が一般的である[8]。また、2000年代までではさほど世間一般に知られていなかったが、中東の兵士から大きく見せるように、カメラの近くまで持ち上げて撮影されたヒヨケムシの写真を起点として徐々に広く知られるようになり、それに連れて様々な不正確な情報もネットなどで拡散された[18][19]。特に有名なものは次の通りに挙げられる。
- 「睡眠中の人を襲う」「上から落ちて人を殺す」
- 「人を追いかける」「ラクダを捕食する」
- 「女性の髪に引き寄せされる」「人の髪や他の動物の毛を切り落とす」
- 「鋏角に噛まれた部分はひどい炎症・痛い腫れ・感覚を失うなどの症状が起きる」
- 「サソリと死闘する」
- 「人並びに早く走れる」「時速48km走れる」「1m高く跳べる」
- 走りは最速時速16kmで、突出した跳躍能力はない[18]。
- 「叫ぶ」
また、クモらしき姿と一部の紛らわしい俗称に相まって、クモ(spider)やサソリ(scorpion)と誤解されてしまうこともある[18][79]。
脚注
[編集]注釈
[編集]- ^ a b 古生代石炭紀ミシシッピ紀(約3億4,000万年前)の Schneidarachne saganii は基盤的なヒヨケムシだった可能性がある。後述参照。
- ^
- 左上:Metasolpuga picta(ヒヨケムシ科)
- 右上:オオヒヨケムシ科の1種
- 左中:Galeodes sp.(サメヒヨケムシ科)
- 右中:ヒトリヒヨケムシ科の1種
- 左下:Gluvia dorsalis(コヒヨケムシ科)
- 右下:Ammotrecha sp.(スナハシリヒヨケムシ科)
- ^ 広義では本群全般、狭義では本群のヒヨケムシ科 Solpugidae のみを指す。
- ^ A: Gylippus rickmersi、B: Galeodes orientalis、C: Eremobates aztecus、D: Pseudocleobis andinus、E: Rhagodes semiflavus、F: Zeria lethalis
- ^ a b Millot & Vachon 1949b では、一般に前体第6節由来とされる metapeltidium が後体第1節(第7体節)由来の背板と解釈される。
- ^ 下咽頭 hypopharynx や下唇 labium ともいうが、他の節足動物における同名の構造とは別器官である。
- ^ 文献により先節/前大脳性の腹板(prosternum, Börner (1902, 1904))、第1体節/中大脳性の腹板(deutosternum, Millot and Vachon 1949, Snodgrass 1948)、第2体節/後大脳性の腹板(tritosternum, Kästner 1931b)と解釈される。
- ^ a b 末端の爪をつ前跗節は通常では肢節扱いされていないが、解剖学上では真の最終肢節である。また、脚の「蹠節」と「跗節」は通常では独立した2肢節扱いされるが、解剖学上では全体的に触肢の跗節に連続相同で、2節(「蹠節」=基跗節、「跗節」=端跗節)に分化した1肢節(=跗節)である。
- ^ 左上:サメヒヨケムシ科、右上:Rhagodes melanus(オオヒヨケムシ科}、中央:Zeria venator(ヒヨケムシ科)、左下:Ammotrecha nigrescens(スナハシリヒヨケムシ科)、右下:Hexisopus abnormis(アナホリヒヨケムシ科)
- ^ 上:Lipophaga michaelseni(アジアヒヨケムシ科 Lipophaginae 亜科)、左下:Eremoperna affinis(ヒトリヒヨケムシ科)、右下:Melanoblossia globiceps(ウモウヒゲヒヨケムシ科 Melanoblossiinae 亜科)
- ^ 鞭毛複合体 flagellar complex 全体を鞭毛 flagellum と扱われる場合もある。
- ^ スナハシリヒヨケムシ科・コヒヨケムシ科・ヒトリヒヨケムシ科・サメヒヨケムシ科・カルシュヒヨケムシ科・コヒヨケムシ科・ヒカラビヒヨケムシ科
- ^
- スナハシリヒヨケムシ科:Oltacola(1種)、Titanopuga(1種)
- コヒヨケムシ科:Gulvia(1種)
- サメヒヨケムシ科:Galeodes(4種)、Othoes(1種)
- ヒトリヒヨケムシ科:Eremobates(6種)、Eremochelis(1種)、Eremopus(1種)
- ヒヨケムシ科:Metasolpuga(1種)
- ^ アジアヒヨケムシ科 Gylippidae、ヒトリヒヨケムシ科 Eremobatidae、カルシュヒヨケムシ科 Karschiidae、サメヒヨケムシ科 Galeodidae、オオヒヨケムシ科 Rhagodidae
- ^ ミナミヒヨケムシ科 Ceromidae、アナホリヒヨケムシ科 Hexisopodidae、ヒヨケムシ科 Solpugidae、コヒヨケムシ科 Daesiidae、ウモウヒゲヒヨケムシ科 Melanoblossidae(Dinorhax除く)、スナハシリヒヨケムシ科 Ammotrechidae、ヒカラビヒヨケムシ科 Mummuciidae
- ^ a b c d e f g h i j k l 特記しない限り Harvey (2003, 2013) と Bird et al. 2015 に基づく。
- ^ Iuri et al. 2021 まででは Mortola は独立属とされ、独自に Mortolinae 亜科に分類された。
- ^ Wharton 1981 では独自に Namibesiinae 亜科に分類される。
- ^ a b ヒカラビヒヨケムシ科の Vempironiella aguilari(3.9 - 5.85mm)が記載される以前では、ウモウヒゲヒヨケムシ科の Lawrencega minuta(5 - 8mm)が知られる中で最小のヒヨケムシであった。
出典
[編集]- ^ a b c d e f g h i j k l Dunlop, Jason A.; Bird, Tharina L.; Brookhart, Jack O.; Bechly, Günter (2015-09-01). “A camel spider from Cretaceous Burmese amber” (英語). Cretaceous Research 56: 265–273. doi:10.1016/j.cretres.2015.05.003. ISSN 0195-6671.
- ^ a b Sundevall, C.J. (1833). Conspectus Arachnidum. Londini Gothorum: Typis Excudit C.F. Berling, Univ. Typogr. 39 pp.
- ^ a b c d e Armas, Luis F. de (2004). “Arácnidos de República Dominicana. Palpigradi, Schizomida, Solifugae y Thelyphonida (Chelicerata: Arachnida)”. Revista ibérica de aracnología (Extra 2): 4–63. ISSN 1576-9518.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay az ba bb bc bd be bf bg bh bi bj bk bl bm bn bo bp bq br bs bt bu bv bw bx by bz ca cb cc cd ce cf cg ch ci cj ck cl cm cn co cp cq cr cs ct cu cv cw Bird, Tharina L.; Wharton, Robert A.; Prendini, Lorenzo (2015-06). “Cheliceral Morphology in Solifugae (Arachnida): Primary Homology, Terminology, and Character Survey”. Bulletin of the American Museum of Natural History 2015 (394): 1-356. doi:10.1206/916.1. ISSN 0003-0090.
- ^ a b c d e f g h i j k Selden, Paul A.; Shear, William A. (1996). “The first Mesozoic Solifugae (Arachnida), from the Cretaceous of Brazil, and a redescription of the Palaeozoic solifuge”. Palaeontology 39: 583–604. ISSN 0031-0239.
- ^ a b c d e f g h i j k l m n o p q r s t “Solifugae (solifuges, solifugids, solpugids)”. www.biodiversityexplorer.info. 2022年5月8日閲覧。
- ^ a b c d e Bittel, Jason. “Camel Spiders Are Fast, Furious and Horrifically Fascinating” (英語). Smithsonian 2018年11月18日閲覧。
- ^ a b c d e Dehghani, R. (2017). “Solpugidophobia in Iran: Real or Illusion”. Journal of Biology and Today's World 6 (3). doi:10.15412/J.JBTW.01060302.
- ^ a b c d e f g h i j k l m n o p q r s t u v Harvey, Mark S (2003). “Catalogue of the Smaller Arachnid Orders of the World” (英語). CSIRO Publishing. doi:10.1071/9780643090071.
- ^ a b c “Introduction: What Are Solifuges?”. www.solpugid.com (2006年). 2018年11月19日閲覧。
- ^ a b “Fossil arachnids” (英語). Deposits Magazine. (2016年8月30日) 2018年11月19日閲覧。
- ^ a b c d e f g h i j k l m Reddick, Kristie Lynn (2009-05-15) (英語). The diversity, distribution and feeding behavior of solifuges (arachnida; solifugae) in Kenya..
- ^ a b c d e f g h i j k l m n o p q r s t Peretti, Alfredo V.; Vrech, David E.; Hebets, Eileen A. (2021-12). “Solifuge (camel spider) reproductive biology: an untapped taxon for exploring sexual selection”. The Journal of Arachnology 49 (3): 299–316. doi:10.1636/JoA-S-20-037. ISSN 0161-8202.
- ^ 『sol』2021年9月11日。2022年5月10日閲覧。
- ^ 『fugere』2021年7月2日。2022年5月10日閲覧。
- ^ a b クモ学―摩訶不思議な八本足の世界. 小野 展嗣. 東海大学出版会. (2002.6). ISBN 4-486-01589-4. OCLC 674859858
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab Klann, Anja Elisabeth (2009). Histology and ultrastructure of solifuges : comparative studies of organ systems of solifuges (Arachnida, Solifugae) with special focus on functional analyses and phylogenetic interpretations.
- ^ a b c d e f g h i j k “Camel Spiders: Facts & Myths”. Live Science. (2014年12月17日) 2018年11月20日閲覧。
- ^ a b c d e “Camel Spider - Facts and Beyond” (英語). Biology Dictionary (2020年7月18日). 2022年5月8日閲覧。
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z Punzo, Fred (1998-04-30) (英語). The Biology of Camel-Spiders: Arachnida, Solifugae. Springer US. ISBN 9780792381556
- ^ “phalangion - 英語 定義、文法、発音、類義語、例文 | Glosbe”. glosbe.com. 2022年5月10日閲覧。
- ^ a b c d Botero-Trujillo, Ricardo (2016). “The smallest known solifuge: Vempironiella aguilari, new genus and species of sun-spider (Solifugae: Mummuciidae) from the coastal desert of Peru”. The Journal of Arachnology 44 (2): 218–226.
- ^ a b c d e f g h i j k Dunlop, Jason A.; Lamsdell, James C. (2017-05-01). “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3): 395–418. doi:10.1016/j.asd.2016.05.002. ISSN 1467-8039.
- ^ a b c d e Lamsdell, James C. (2013-01). “Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura: Re-evaluating the Monophyly of Xiphosura” (英語). Zoological Journal of the Linnean Society 167 (1): 1–27. doi:10.1111/j.1096-3642.2012.00874.x.
- ^ a b Lankester, Edwin Ray (1911), Encyclopædia Britannica, Volume 2: Arthropoda, 2
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai ROEWER, C.F. 1932-1934. Solifuga, Palpigrada. In E.G. Bronn (editor), Klassen und Ordnungen des Tierreichs. 5: Arthropoda. IV: Arachnoidea und kleinere ihnen nahegestellte Gruppen. Vol. 5 (IV) (4) (1–5): 481– 723. Leip
- ^ a b c Dunlop, Jason A.; Krüger, Jessica; Alberti, Gerd (2012-07-31). “The sejugal furrow in camel spiders and acariform mites” (英語). Arachnologische Mitteilungen 43: 29–36. doi:10.5431/aramit4303. ISSN 1018-4171.
- ^ Miether, Sebastian T.; Dunlop, Jason A. (2016-07). “Lateral eye evolution in the arachnids”. Arachnology 17 (2): 103–119. doi:10.13156/arac.2006.17.2.103. ISSN 2050-9928.
- ^ a b c d e f Yamasaki, Takeshi; Henriques, Sérgio; Thi Hong Phung, , Luong; Hoang, Quang Duy (2018-12-01). “Redescription of the sole species of the enigmatic solifuge genus Dinorhax Simon, 1879 (Solifugae: Melanoblossiidae) in Southeast Asia”. Journal of Arachnology 46: 498–506. doi:10.1636/JoA-S-17-090.1.
- ^ a b c d Runge, Jens; Wirkner, Christian S. (2020-10-26). “Evolutionary and functional substitution of extrinsic musculature in Solifugae (Arachnida)” (英語). Journal of Morphology 281 (12): 1524–1533. doi:10.1002/jmor.21260. ISSN 0362-2525.
- ^ a b c d e f g h i j k Franz-Guess, Sandra; Klußmann-Fricke, Bastian-Jesper; Wirkner, Christian S.; Prendini, Lorenzo; Starck, J. Matthias (2016-09-01). “Morphology of the tracheal system of camel spiders (Chelicerata: Solifugae) based on micro-CT and 3D-reconstruction in exemplar species from three families” (英語). Arthropod Structure & Development 45 (5): 440–451. doi:10.1016/j.asd.2016.08.004. ISSN 1467-8039.
- ^ a b c d e f g Dunlop (2000). “The epistomo-labral plate and lateral lips in solifuges, pseudoscorpions and mites”. Ekológia, Bratislava 19: 67–78.
- ^ a b Shultz, Jeffrey W. (1990-03). “EVOLUTIONARY MORPHOLOGY AND PHYLOGENY OF ARACHNIDA”. Cladistics 6 (1): 1–38. doi:10.1111/j.1096-0031.1990.tb00523.x. ISSN 0748-3007.
- ^ a b c Starck, J. Matthias; Belojević, Jelena; Brozio, Jason; Mehnert, Lisa (2022-03-01). “Comparative anatomy of the rostrosoma of Solifugae, Pseudoscorpiones and Acari” (英語). Zoomorphology 141 (1): 57–80. doi:10.1007/s00435-021-00551-3. ISSN 1432-234X.
- ^ a b Hrušková-Martišová, Martina; Pekár, Stano; Gromov, Alexandr (2008-07-22). “Analysis of the Stridulation in Solifuges (Arachnida: Solifugae)” (英語). Journal of Insect Behavior 21 (5): 440. doi:10.1007/s10905-008-9141-4. ISSN 1572-8889.
- ^ van der Meijden, Arie; Langer, Franz; Boistel, Renaud; Vagovic, Patrik; Heethoff, Michael (2012-10-01). “Functional morphology and bite performance of raptorial chelicerae of camel spiders (Solifugae)”. Journal of Experimental Biology 215 (19): 3411–3418. doi:10.1242/jeb.072926. ISSN 0022-0949.
- ^ a b Cushing, Paula E.; Brookhart, Jack O.; Kleebe, Hans-Joachim; Zito, Gary; Payne, Peter (2005-10-01). “The suctorial organ of the Solifugae (Arachnida, Solifugae)” (英語). Arthropod Structure & Development 34 (4): 397–406. doi:10.1016/j.asd.2005.02.002. ISSN 1467-8039.
- ^ Klann, A. E.; Gromov, A. V.; Cushing, P. E.; Peretti, A. V.; Alberti, G. (2008-01-01). “The anatomy and ultrastructure of the suctorial organ of Solifugae (Arachnida)” (英語). Arthropod Structure & Development 37 (1): 3–12. doi:10.1016/j.asd.2007.04.001. ISSN 1467-8039.
- ^ Willemart, Rodrigo; Santer, Roger; Spence, Andrew; Hebets, Eileen (2011-01-01). “A sticky situation: Solifugids (Arachnida, Solifugae) use adhesive organs on their pedipalps for prey capture”. Eileen Hebets Publications.
- ^ a b c d e f g SHULTZ, JEFFREY W. (1989-09-01). “Morphology of locomotor appendages in Arachnida: evolutionary trends and phylogenetic implications”. Zoological Journal of the Linnean Society 97 (1): 1–56. doi:10.1111/j.1096-3642.1989.tb00552.x. ISSN 0024-4082.
- ^ a b Sensenig, Andrew T.; Shultz, Jeffrey W. (2003-02-15). “Mechanics of cuticular elastic energy storage in leg joints lacking extensor muscles in arachnids” (英語). Journal of Experimental Biology 206 (4): 771–784. doi:10.1242/jeb.00182. ISSN 0022-0949. PMID 12517993.
- ^ a b c d e f g h i j k l m n o p q “Key to Families”. www.solpugid.com. 2018年11月18日閲覧。
- ^ a b c d Klann, Anja E.; Bird, Tharina L.; Talarico, Giovanni (2011). “Ultrastructural characterization of Hexisopus psammophilus (Arachnida: Solifugae: Hexisopodidae) spermatozoa in comparison to other solifuge spermatozoal traits”. The Journal of arachnology 39 (2): 280–286. doi:10.1636/cb10-94.1. ISSN 0161-8202.
- ^ a b c d Sombke, Andy; Klann, Anja E.; Lipke, Elisabeth; Wolf, Harald (2019-08-02). “Primary processing neuropils associated with the malleoli of camel spiders (Arachnida, Solifugae): a re-evaluation of axonal pathways”. Zoological Letters 5 (1): 26. doi:10.1186/s40851-019-0137-z. ISSN 2056-306X. PMC 6679463. PMID 31388441.
- ^ a b c d e f g h i j k l m n Muma, Martin H. (1967). “Basic Behavior of North American Solpugida”. The Florida Entomologist 50 (2): 115–123. doi:10.2307/3493620. ISSN 0015-4040.
- ^ a b c van der Hammen, L. (1986-01-01). “Comparative studies in Chelicerata IV. Apatellata, Arachnida, Scorpionida, Xiphosura” (英語). Zoologische Verhandelingen 226 (1): 1–52.
- ^ a b Schultz, Jeffrey Walden (1990) (英語). Evolutionary morphology of the locomotor apparatus in arachnida.
- ^ a b c McLean, Callum J.; Garwood, Russell J.; Brassey, Charlotte A. (2018-11-06). “Sexual dimorphism in the Arachnid orders” (英語). PeerJ 6: e5751. doi:10.7717/peerj.5751. ISSN 2167-8359.
- ^ a b Ryan Jones, R.; Cushing, Paula E. (2021-11-01). “An assessment of function, intraspecific variation, and taxonomic reliability of eremobatid ctenidia (Arachnida: Solifugae)” (英語). Zoologischer Anzeiger 295: 43–54. doi:10.1016/j.jcz.2021.09.002. ISSN 0044-5231.
- ^ KARATAŞ, AYŞEGÜL; UÇAK, MÜNİR (2013-01-01). “A new Barrussus Roewer, 1928 (Solifugae: Karschiidae) from southern Turkey”. Turkish Journal of Zoology 37 (5): 594–600. doi:10.3906/zoo-1202-16. ISSN 1300-0179.
- ^ a b Alberti, G.; Peretti, A. V. (2002-08). [0268:FSOMGS2.0.CO;2.full “FINE STRUCTURE OF MALE GENITAL SYSTEM AND SPERM IN SOLIFUGAE DOES NOT SUPPORT A SISTER-GROUP RELATIONSHIP WITH PSEUDOSCORPIONES (ARACHNIDA)”]. The Journal of Arachnology 30 (2): 268–274. doi:10.1636/0161-8202(2002)030[0268:FSOMGS]2.0.CO;2. ISSN 0161-8202.
- ^ a b Klann, A. E.; Bird, T.; Peretti, A. V.; Gromov, A. V.; Alberti, G. (2009-04-01). “Ultrastructure of spermatozoa of solifuges (Arachnida, Solifugae): Possible characters for their phylogeny?” (英語). Tissue and Cell 41 (2): 91–103. doi:10.1016/j.tice.2008.07.003. ISSN 0040-8166.
- ^ a b c Ballesteros, Jesús A.; Santibáñez López, Carlos E.; Kováč, Ľubomír; Gavish-Regev, Efrat; Sharma, Prashant P. (2019-12-18). “Ordered phylogenomic subsampling enables diagnosis of systematic errors in the placement of the enigmatic arachnid order Palpigradi”. Proceedings of the Royal Society B: Biological Sciences 286 (1917): 20192426. doi:10.1098/rspb.2019.2426. PMC 6939912. PMID 31847768.
- ^ a b c d e f g h i j k l m n Belozerov, V. N. (2013-11). “Seasonal aspects of the life cycle of solifuges (Arachnida, Solifugae) as compared with pseudoscorpions (Arachnida, Pseudoscorpiones)” (英語). Entomological Review 93 (8): 1050–1072. doi:10.1134/S0013873813080113. ISSN 0013-8738.
- ^ Polis, Gary A.; McCormick, Sharon J. (1986-12). “Scorpions, spiders and solpugids: predation and competition among distantly related taxa”. Oecologia 71 (1): 111–116. doi:10.1007/bf00377328. ISSN 0029-8549.
- ^ “Biology, Behavior, and Ecology”. www.solpugid.com. 2022年5月8日閲覧。
- ^ a b c d e f g h i j Robert A. Wharton (1986). “Biology of the Diurnal Metasolpuga picta (Kraepelin) (Solifugae, Solpugidae) Compared with That of Nocturnal Species”. The Journal of Arachnology Vol. 14, No. 3: pp. 363-383.
- ^ “Egyptian giant solpugid (camel spider) Galeodes arabs”. National Geographic. June 10, 2011閲覧。
- ^ Brownell, Philip H.; Farley, Roger D. (1974-01-01). “The organization of the malleolar sensory system in the solpugid, Chanbria sp.” (英語). Tissue and Cell 6 (3): 471–485. doi:10.1016/0040-8166(74)90039-1. ISSN 0040-8166.
- ^ a b c Graham, Matthew R.; Pinto, Michelina B.; Cushing, Paula E. (2019-09-16). “A test of the light attraction hypothesis in camel spiders of the Mojave Desert (Arachnida: Solifugae)”. The Journal of Arachnology 47 (2): 293. doi:10.1636/JoA-S-18-077. ISSN 0161-8202.
- ^ a b “Prey”. www.solpugid.com. 2022年5月8日閲覧。
- ^ a b c d Wharton, Robert A.; Reddick, Kristie L. (2014-09-02). “Solifuges (Arachnida: Solifugae) as predators and prey”. Transactions of the Royal Society of South Africa 69 (3): 213–216. doi:10.1080/0035919X.2014.961045. ISSN 0035-919X.
- ^ a b c d e f g Muma, Martin H. (1966). “Feeding Behavior of North American Solpugida (Arachnida)”. The Florida Entomologist 49 (3): 199–216. doi:10.2307/3493444. ISSN 0015-4040.
- ^ “Scavenging Behavior”. www.solpugid.com. 2022年5月8日閲覧。
- ^ “Foraging Behavior”. www.solpugid.com. 2022年5月8日閲覧。
- ^ Hrušková-Martišová, Martina; Pekár, Stano; Gromov, Alexander (2007-12). “Biology Of Galeodes caspius subfuscus (Solifugae, Galeodidae)”. The Journal of Arachnology 35 (3): 546–550. doi:10.1636/JOA/06-64.1. ISSN 0161-8202.
- ^ a b c “Predators”. www.solpugid.com. 2022年5月8日閲覧。
- ^ a b c d e Hrušková‐Martišová, M.; Pekár, S.; Bilde, T. (2010-10). “Coercive copulation in two sexually cannibalistic camel‐spider species (Arachnida: Solifugae)” (英語). Journal of Zoology 282 (2): 91–99. doi:10.1111/j.1469-7998.2010.00718.x. ISSN 0952-8369.
- ^ Hrušková-Martišová, Martina; Pekár, Stano; Gromov, Alexandr (2008-07-22). “Analysis of the Stridulation in Solifuges (Arachnida: Solifugae)” (英語). Journal of Insect Behavior 21 (5): 440–449. doi:10.1007/s10905-008-9141-4. ISSN 0892-7553.
- ^ a b “Biology, Behavior, and Ecology (Mimicry)”. www.solpugid.com. 2018年11月18日閲覧。
- ^ a b c d Rowsell, Jenny; Cushing, Paula E. (2020-03). “Mating behaviour of Eremobates pallipes (Say, 1823) (Arachnida: Solifugae: Eremobatidae)”. Arachnology 18 (4): 399–408. doi:10.13156/arac.2020.18.4.399. ISSN 2050-9928.
- ^ a b “Courtship and Mating of Solifuges”. www.solifugae.info. 2018年11月18日閲覧。
- ^ a b c “Courtship and Mating”. www.solpugid.com. 2022年5月8日閲覧。
- ^ “The Naming of Parts | Natural History Magazine” (英語). www.naturalhistorymag.com. 2018年11月18日閲覧。
- ^ Peretti, Alfredo V.; Vrech, David E.; Hebets, Eileen A. (2021-12). “Solifuge (camel spider) reproductive biology: an untapped taxon for exploring sexual selection”. The Journal of Arachnology 49 (3): 299–316. doi:10.1636/JoA-S-20-037. ISSN 0161-8202.
- ^ G. Schmidt (1993). Giftige und gefährliche Spinnentiere (in German). Westarp Wissenschaften. ISBN 3-89432-405-8.
- ^ a b c “Life History”. www.solpugid.com. 2022年5月8日閲覧。
- ^ Erdek, Melek (2021-06-30). “An Evaluation on the Last Ecdysis in Galeodids (Galeodidae, Solifugae, Arachnida)” (英語). Commagene Journal of Biology 5 (1): 90–93. doi:10.31594/commagene.906878.
- ^ a b “世界三大奇虫:クモなのに脚10本「ヒヨケムシ」日本で初”. 毎日新聞. 2019年9月28日閲覧。
- ^ Hayashi; Wheeler, W.; Platnick, N. I. (1992). “Molecular phylogenetics of chelicerates with a special emphasis on basal araneomorphs”. American Arachnology 46: 9.
- ^ a b Wheeler, Ward C.; Hayashi, Cheryl Y. (1998). “The Phylogeny of the Extant Chelicerate Orders” (英語). Cladistics 14 (2): 173–192. doi:10.1111/j.1096-0031.1998.tb00331.x. ISSN 1096-0031.
- ^ Giribet, G (2002-02). “Phylogeny and Systematic Position of Opiliones: A Combined Analysis of Chelicerate Relationships Using Morphological and Molecular Data” (英語). Cladistics 18 (1): 5–70. doi:10.1006/clad.2001.0185.
- ^ SHULTZ, JEFFREY W. (2007-06-01). “A phylogenetic analysis of the arachnid orders based on morphological characters”. Zoological Journal of the Linnean Society 150 (2): 221–265. doi:10.1111/j.1096-3642.2007.00284.x. ISSN 0024-4082.
- ^ Giribet, Gonzalo; Edgecombe, Gregory D. (2012). “Reevaluating the arthropod tree of life”. Annual Review of Entomology 57: 167–186. doi:10.1146/annurev-ento-120710-100659. ISSN 1545-4487. PMID 21910637.
- ^ a b c d e Cushing, Paula E.; Graham, Matthew R.; Prendini, Lorenzo; Brookhart, Jack O. (2015-11-01). “A multilocus molecular phylogeny of the endemic North American camel spider family Eremobatidae (Arachnida: Solifugae)” (英語). Molecular Phylogenetics and Evolution 92: 280–293. doi:10.1016/j.ympev.2015.07.001. ISSN 1055-7903.
- ^ a b c Dunlop, Jason; Garwood, Russell J. (2014-11-13). “Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders” (英語). PeerJ 2: e641. doi:10.7717/peerj.641. ISSN 2167-8359.
- ^ Weygoldt, P.; Paulus, H. F. (1979). “Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata1 I. Morphologische Untersuchungen” (英語). Journal of Zoological Systematics and Evolutionary Research 17 (2): 85–116. doi:10.1111/j.1439-0469.1979.tb00694.x.
- ^ Regier, Jerome C.; Shultz, Jeffrey W.; Zwick, Andreas; Hussey, April; Ball, Bernard; Wetzer, Regina; Martin, Joel W.; Cunningham, Clifford W. (2010-02). “Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences” (英語). Nature 463 (7284): 1079–1083. doi:10.1038/nature08742. ISSN 1476-4687.
- ^ Howard, Richard J.; Puttick, Mark N.; Edgecombe, Gregory D.; Lozano-Fernandez, Jesus (2020-11-01). “Arachnid monophyly: Morphological, palaeontological and molecular support for a single terrestrialization within Chelicerata” (英語). Arthropod Structure & Development 59: 100997. doi:10.1016/j.asd.2020.100997. ISSN 1467-8039.
- ^ a b Pepato, Almir R.; da Rocha, Carlos EF; Dunlop, Jason A. (2010-08-02). “Phylogenetic position of the acariform mites: sensitivity to homology assessment under total evidence”. BMC Evolutionary Biology 10 (1): 235. doi:10.1186/1471-2148-10-235. ISSN 1471-2148. PMC 2933639. PMID 20678229.
- ^ Ballesteros, Jesús A; Santibáñez-López, Carlos E; Baker, Caitlin M; Benavides, Ligia R; Cunha, Tauana J; Gainett, Guilherme; Ontano, Andrew Z; Setton, Emily V W et al. (2022-02-01). “Comprehensive Species Sampling and Sophisticated Algorithmic Approaches Refute the Monophyly of Arachnida”. Molecular Biology and Evolution 39 (2): msac021. doi:10.1093/molbev/msac021. ISSN 1537-1719.
- ^ Sharma, Prashant P.; Kaluziak, Stefan T.; Pérez-Porro, Alicia R.; González, Vanessa L.; Hormiga, Gustavo; Wheeler, Ward C.; Giribet, Gonzalo (2014-08-08). “Phylogenomic Interrogation of Arachnida Reveals Systemic Conflicts in Phylogenetic Signal” (英語). Molecular Biology and Evolution 31 (11): 2963–2984. doi:10.1093/molbev/msu235. ISSN 1537-1719.
- ^ Lozano-Fernandez, Jesus; Tanner, Alastair R.; Giacomelli, Mattia; Carton, Robert; Vinther, Jakob; Edgecombe, Gregory D.; Pisani, Davide (2019-05-24). “Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida” (英語). Nature Communications 10 (1): 2295. doi:10.1038/s41467-019-10244-7. ISSN 2041-1723.
- ^ a b c d e f g h i j k l m n o p q r s t u Dunlop, J. A., Penney, D. & Jekel, D. 2020. A summary list of fossil spiders and their relatives. In World Spider Catalog. Natural History Museum Bern, online at http://wsc.nmbe.ch, version 20.5
- ^ a b Maddahi, Hassan; Khazanehdari, Mahsa; Aliabadian, Mansour; Kami, Haji Gholi; Mirshamsi, Amin; Mirshamsi, Omid (2016-10-28). “Mitochondrial DNA phylogeny of camel spiders (Arachnida: Solifugae) from Iran”. Mitochondrial DNA Part A 28 (6): 909–919. doi:10.1080/24701394.2016.1209194. ISSN 2470-1394. PMID 27822966.
- ^ a b Iuri, Hernán A.; Iglesias, Mónica S. (2022-02-04). “The genus Pseudocleobis Pocock, 1900 (Solifugae: Ammotrechidae) in transitional Patagonia-Monte deserts, with descriptions of two new species” (英語). Zootaxa 5094 (3): 435–460. doi:10.11646/zootaxa.5094.3.4. ISSN 1175-5334.
- ^ a b c Erdek, Melek (2021-06-23). “A new species of the solifuge genus Galeodes Olivier, 1791 from southeastern Turkey (Solifugae, Galeodidae)” (英語). Zootaxa 4991 (1): 116–130. doi:10.11646/zootaxa.4991.1.5. ISSN 1175-5334.
- ^ a b c Souza, Maysa F. V. R.; Ferreira, Rodrigo L.; Carvalho, Leonardo S. (2021-11-05). “A new species of the genus Gaucha Mello-Leitão, 1924 from Minas Gerais, Brazil (Solifugae, Mummuciidae)” (英語). Zootaxa 5061 (3): 559–572. doi:10.11646/zootaxa.5061.3.9. ISSN 1175-5334.
- ^ Kulkarni, Siddharth S.; Steiner, Hugh G.; Garcia, Erika L.; Iuri, Hernán; Jones, R. Ryan; Ballesteros, Jesús A.; Gainett, Guilherme; Graham, Matthew R. et al. (2023-09). “Neglected no longer: Phylogenomic resolution of higher-level relationships in Solifugae”. iScience 26 (9): 107684. doi:10.1016/j.isci.2023.107684. ISSN 2589-0042. PMC PMC10484990. PMID 37694155.
- ^ a b c d Iuri, Hernán A.; Ramírez, Martín J.; Mattoni, Camilo I.; Ojanguren-Affilastro, Andrés A. (2021-11-01). “Revision and cladistic analysis of subfamily Nothopuginae (Solifugae, Ammotrechidae)” (英語). Zoologischer Anzeiger 295: 126–155. doi:10.1016/j.jcz.2021.10.001. ISSN 0044-5231.
- ^ a b Poinar, George O.; Santiago-Blay, J. A. (1989). “A Fossil Solpugid, Happlodontus proterus, New Genus, New Species (Arachnida: Solpugida) from Dominican Amber”. Journal of the New York Entomological Society 97: 125–132. ISSN 0028-7199.
- ^ Muma, Martin H. (Martin Hammond); Peña G., Luis E.; Schlinger, Evert I. (Evert Irving); Irwin, Michael E. (1971) (英語). The solpugids (Arachnida, Solpugida) of Chile, with descriptions of a new family, new genera, and new species. American Museum novitates ; no. 2476.
- ^ Maury, E.A. 1985. Nota sobre los géneros Namibesia y Syndaesia (Solifugae, Daesiidae). Aracnología 4: 1-6.
- ^ “Family Eremobatidae”. www.solpugid.com. 2018年11月18日閲覧。
- ^ Muma, M.H. 1970a. A synoptic review of North American, Central American, and West Indian Solpugida (Arthropoda, Arachnida). Arthropods of Florida and Neighboring Land Areas 5: 1–62.
- ^ “Galeodidae”. www.solpugid.com. 2022年5月14日閲覧。
- ^ Bird, Tharina L.; Wharton, Robert A.; Prendini, Lorenzo (2015-06). “Cheliceral Morphology in Solifugae (Arachnida): Primary Homology, Terminology, and Character Survey”. Bulletin of the American Museum of Natural History 2015 (394): 1–356. doi:10.1206/916.1. ISSN 0003-0090.
- ^ “The natural history of teddy-bear solifugids: cuddly wonders of the desert” (英語). Arthropod Ecology. (2015年10月16日) 2018年11月18日閲覧。
- ^ a b c d Bird, Tharina L.; Wharton, Robert A. (2015-08). “Description of a New Solifuge Melanoblossia ansie sp. n. (Solifugae, Melanoblossiidae) with Notes on the Setiform Flagellar Complex of Melanoblossiinae Roewer, 1933”. African Invertebrates 56 (2): 515–525. doi:10.5733/afin.056.0218. ISSN 1681-5556.
- ^ a b Bartel, Christian; Dunlop, Jason A.; Bird, Tharina L. (2016-11). “The Second Camel Spider (Arachnida, Solifugae) from Burmese Amber”. Arachnology 17 (3): 161–164. doi:10.13156/arac.2006.17.3.161. ISSN 2050-9928.
- ^ Botero-Trujillo, Ricardo; Lagos-Silnik, Susana; Fernández-Campón, Florencia (2019-12). “Curanahuel aconcagua, a new genus and species of sun-spider (Solifugae: Mummuciidae) from the Cuyan High Andean biogeographic province of Argentina”. The Journal of Arachnology 47 (3): 351–359. doi:10.1636/0161-8202-47.3.351. ISSN 0161-8202.
- ^ Trujillo, Ricardo Botero; Ott, R.; Carvalho, L. S. (2017). “Systematic revision and phylogeny of the South American sun-spider genus Gaucha Mello-Leitão (Solifugae: Mummuciidae), with description of four new species and two new generic synonymies” (英語). undefined.
- ^ Cossios, E. Daniel (2020-08-30). “Description of a new Mummucina species (Solifugae: Mummuciidae) from Peru”. Revista Peruana de Biología 27 (3): 283–288. doi:10.15381/rpb.v27i3.18642. ISSN 1727-9933.
- ^ a b “The Arachnid Order Solifugae (Rhagodidae)”. www.solpugid.com. 2018年11月18日閲覧。
- ^ a b “Fossil History”. www.solpugid.com. 2018年11月19日閲覧。
- ^ a b Dunlop, J.; Klann, A. (2009). “A second camel spider (Arachnida: Solifugae) from Baltic amber” (英語). Acta Geologica Polonica Vol. 59 (no. 1): 39–44. ISSN 0001-5709.
- ^ Dunlop, Jason; Rößler, Ronny (2003-04-28). “An enigmatic, solifuge-like fossil arachnid from the Lower Carboniferous of Kamienna Göra (Intra-Sudetic Basin), Poland”. Paläontologische Zeitschrift 77: 389–400. doi:10.1007/BF03006949.
- ^ Dunlop, J. A. & Martill, D. M. 2004. Four additional specimens of the fossil camel spider Cratosolpuga wunderlichi Selden 1996 (Arachnida: Solifugae) from the Lower Cretaceous Crato Formation of Brazil. Revista Ibérica de Aracnología 9: 143-156.
- ^ Dunlop, Jason A.; Wunderlich, Jörg; Poinar, George O. (2003-09). “The first fossil opilioacariform mite (Acari: Opilioacariformes) and the first Baltic amber camel spider (Solifugae)”. Transactions of the Royal Society of Edinburgh: Earth Sciences 94 (03): 261–273. doi:10.1017/s0263593300000663. ISSN 0263-5933.
- ^ a b Cushing, Paula E.; González-Santillán, Edmundo (2018-08). “Capturing the elusive camel spider (Arachnida: Solifugae): effective methods for attracting and capturing solifuges”. The Journal of Arachnology 46 (2): 384–387. doi:10.1636/JoA-S-17-067.1. ISSN 0161-8202.
- ^ a b c d e Kassiri; Mazaheri-Tehrani; Yaselyani; Akbarzadeh (2019). A preliminary study on fauna of medical important solpugid (Chelicerata: Arachnida: Solifugae) in Kashan City, Central Iran.. doi:10.4066/biomedicalresearch.30-18-1191.
参考文献
[編集]- 内田亨監修 『動物系統分類学』第7巻(中A)「真正蜘蛛類」、中山書店。
関連項目
[編集]外部リンク
[編集]- ヒヨケムシ総説:
- Solifuges of the World | Field Guide and Catalogues | www.museum.wa.gov.au - ヒヨケムシのフィールドガイドと目録(2011年末までの情報に基づく)
- Camel Spiders: Facts & Myths | Live Science - ヒヨケムシに関するファクトチェック