
サソリ
| サソリ | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 様々なサソリ[注釈 1]
| ||||||||||||||||||
| 分類 | ||||||||||||||||||
| ||||||||||||||||||
| 学名 | ||||||||||||||||||
| Scorpiones C. L. Koch, 1837 | ||||||||||||||||||
| 英名 | ||||||||||||||||||
| Scorpion | ||||||||||||||||||
| 下位分類群 | ||||||||||||||||||
|
本文参照
|
サソリ(さそり、蠍、蝎、サソリ類、英:scorpion、学名:Scorpiones)は、鋏角亜門クモガタ綱に分類される節足動物の分類群の一つ。分類学上はサソリ目とされる。体の前に鋏型の触肢、後ろに毒針を持つ捕食者である。
1,700以上の種が記載され、最古の化石記録は4億3,000万年前のシルル紀まで遡る。有毒生物として広く知られるが、人間の命にかかわる毒を持つものはごく一部の種類に過ぎない。
日本にはマダラサソリ及びヤエヤマサソリの2種が産する。また、日本ではウシコロシサソリ科全種が特定外来生物に指定されている。
形態
[編集]
.png)
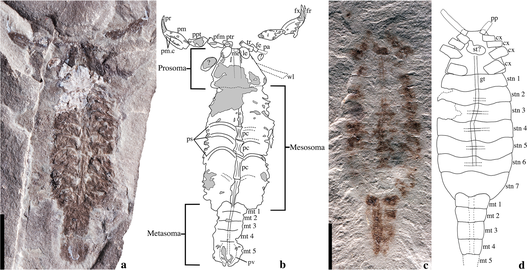
体は縦長く、短い前体(頭胸部)と長い後体(腹部)で構成されており、その間はくびれていない。後体は更に丈夫な中体(前腹部)と細長い終体(後腹部)に分化される[3]。鋏型の触肢・尻尾のように特化した終体・鈎状の毒針に特化した尾節が特徴である[3]。
前体
[編集]
-
 サソリの顔。中眼・側眼・鋏角・触肢の一部が映る
サソリの顔。中眼・側眼・鋏角・触肢の一部が映る -

_chelicerae.JPG)
.jpg)
前体(prosoma、頭胸部 cephalothorax ともいうが、頭部そのものに相当[4])は先節と直後6つの体節から癒合した合体節であり、後体より遥かに短く、背面は1枚の背甲(carapace, prosomal dorsal shield)に覆われる[3]。背甲は正中線に1対の中眼(median eyes)と両前端に側眼(lateral eyes)をもつ。現生群では中眼は背甲の中央付近に位置し、側眼は通常3対(最少2対、最多5対[5])の単眼であるが、基盤的な化石種では中眼は背甲の前端に位置し、側眼は複眼的である[5][6]。なお、全く眼を欠く洞穴性の種類もある[7][8]。
他のクモガタ類と同様、前体は鋏角1対・触肢1対・脚4対という計6対の付属肢(関節肢)をもつ[3]。短い鋏角(chelicerae)は3節からなり、短縮した基部の1節(柄部)は常に背甲の下に隠れ、先端の2節は鋏をなしている。触肢(pedipalp)は鋏角の下から大きく張り出し、6節からなり、先端の2節が発達した鋏を構成する。触肢の直後は7節に分かれた4対の脚があり、後方ほど長くなる。特に第3-4脚の基節は後体の腹面まで長く伸びるため、後ろ脚が後体に備わるようにも見える。目立たない上唇に覆われた口は鋏角と触肢の間に開く[9]。第1-2脚の基節から顎葉(coxapophyses、apophysis)という腹側から口を覆う部分が突き出し、鋏角と触肢基節に併せて「stomotheca」というザトウムシと似た口器をなしている[6][10](基盤的な化石種では顎葉を欠くため stomotheca をなしていない[11][6])。脚の間は1枚の小さな腹板(sternum)が占めており、通常は左右が第3-4脚の基節、前端が第2脚の基節に隣接するが、基盤的な化石種では腹板が縦長く、第1脚の基節まで隣接した場合がある[3][6]。なお、この腹板に関しては、実は前体に由来する腹板ではなく、直後の後体第1節の付属肢から変化したものではないかという説がある[3][12]。
後体
[編集]
後体(opisthosoma、腹部 abdomen)は縦長く、13節と尾節からなるが、その第1節は胚発育の度に退化消失するため、外見上では12節に見える。幅広い前半部は中体(前腹部)、尻尾のような細長い後半部は終体(後腹部)に分化される[3]。
中体
[編集]

-
 Graemeloweus maidu の前体と後体の境目、雌(左)と雄(右)、腹側から。第3-4脚の基節・前体と中体の腹板・生殖口蓋・櫛状板など構造が映る
Graemeloweus maidu の前体と後体の境目、雌(左)と雄(右)、腹側から。第3-4脚の基節・前体と中体の腹板・生殖口蓋・櫛状板など構造が映る
_and_male_sternopectinal_regions.jpg)
中体(mesosoma、前腹部 preabdomen)は丈夫で、後体の前8節を含む。ただし第1節は退化消失し、胚発生の一時期のみに見られる[3][12]。残り7節は外見上で顕著に見られ、外骨格は腹背で背板と腹板に分かれている。ただし第2・3節の腹側は通常では発達した腹板をもたず、代わりにそれぞれ生殖口蓋(genital operculum)という生殖口を覆う板状構造と、櫛状板(ペクチン、pectine)という整髪用の櫛の造形に似た感覚器をもつ[3]。なお、一部の基盤的な化石種では櫛状板の所に発達した腹板をもつ[6][13][14][12]。生殖口蓋は前後で前体の腹板と櫛状板の基部に隣接し、これらの左右が第4脚の長い基節に囲まれる。次の5節のうち前4節の腹面は書肺(book lung)という呼吸器官が1節に1対ずつ配置され、気門を介して空気を出入れされる。生殖口蓋、櫛状板と書肺(および書肺を覆い被さった腹板らしき外骨格)は、いずれも付属肢由来の器官であると考えられる[3][12][15]。
終体
[編集]

.jpeg)
終体(metasoma、後腹部 postabdomen)は細長く、最後5節の体節と尾節からなる[3]。この部分全体が一般にサソリの「尾」と呼ばれるが、実際には特化した後体(腹部)の後半部であるため[3]、形態学や解剖学上の「尾」に該当する部位ではない(真の「尾」は末端の尾節のみに当たる)。5節の外骨格は円筒状で数対の隆起線が走り、可動範囲は広く、通常では上向きに前へ反り曲がる。末端は短い尾節があり、基部は膨らんで毒線を備わり、先端は尖って鈎状の毒針となる[3]。肛門は他の尾節のある鋏角類と同様、尾節と最後の体節の境目の腹側にあるが、終体の反り曲がりの姿勢によってこの肛門は往々にして上向きに露出している。尾節直前の終体第5節は通常では前4節より顕著に長く特化したが、基盤的な化石種ではその長さが往々にして直前の節を超えない[11]。
雌雄
[編集]成体の雌雄の見分け方として、腹部にある櫛状板(ペクチン)が大きい方が往々にして雄であることが挙げられる。
また、雌の方が体が全般的に大きく、太っているが、雄の方は雌を交尾の婚姻ダンスの際に、雌を押さえつけておくために、雌よりも鋏が大きい場合もある。
生態
[編集].jpg)
歩く時は終体を曲げて体の上の前方にのばす。餌を獲った時には、鋏型の触肢で固定した餌に尾部の針を刺し、毒液を注入し、鋏角で獲物を小さくちぎって体液を飲み込む。
肉食で、昆虫や他の節足動物、大型種ではネズミ等の小動物を捕食した例もある。それほど大食いではなく、絶食に耐えるものが多い。
主に夜行性で、昼間は岩の下や土の中、何かの隙間にいることが多い。元来活動はあまり活発ではなく、じっとして獲物が通るのを待っている。
天敵
[編集]昆虫などをエサにするサソリだが、実はサソリ自身にも多くの天敵が存在する。それらの捕食動物相手には、毒針と鋏を振るって応戦するが、相手によっては毒に耐性を持っている場合もあり、自分より大きな動物相手には一方的に捕食されてしまうケースが多い。
サソリの天敵はイタチやジャコウネコ科などの肉食性哺乳類や、鳥類、爬虫類、他に同じサソリや、肉食性の昆虫類にオオヅチグモ類やムカデ類など、他の節足動物にも捕食される。
サソリが一般に暗闇や物陰を好むのも、こういった多くの天敵から逃れる手段ではないかと考えられる。
生殖
[編集]

サソリ類の配偶行動は、婚姻ダンスとして知られる。雌雄が互いの触肢をつかみ合って、前後左右に動き、種によってはそれが数時間以上も続けられる。最終的に、雄は精包を地上に置き、そこへ雌を誘導し、雌はその精包を生殖口から取り入れることで、配偶行動は完了する。
サソリ類は、卵胎生と胎生の種に分けられ、雌親はサソリの形の幼生を産む。生まれた幼生は雌親の体の上に登り、その背中でしばらくの時間を過ごすが、一週間か10日ほどで独立し、自立生活に入る。
ヤエヤマサソリは雌性産生単為生殖することが分かっているが、個体数は少ないものの雄も存在する。その他10ほどの種で単為生殖が知られる。
サソリの婚姻行動は相性の悪い相手であれば、お互いに刺しあってどちらか一方を殺してしまったり、雌が雄を一方的に食べてしまったりするような行動をとってしまうケースもある。
ファーブルはその行動を観察して、サソリはカマキリやクモのように、交尾後に雌が雄を捕食してしまうと思ってしまったが、これは狭い飼育ケージ内での観察であり、野外においての交尾後の共食いは少ないのではないかといわれている。
分布
[編集]サソリ類は世界に多く分布しており、種数は1750種類を超える。基本的には暖かいところに多く、熱帯地方が分布の中心ではあるが、かなり寒い地方まで分布している種もある。日本では、南西諸島に2種が分布するだけだが、アジア大陸では、北朝鮮、内モンゴルにまで分布がある。湿潤な気候に生息する種もあるが、砂漠に生息する種もあり、適応範囲は広い。ヨーロッパでは地中海周辺地域に生息する。人間の生活範囲に生息するものもあり、それらの生活圏内に住む住人は、かならず靴を履く前に、中にサソリが入っていないか調べると言われる。このような種は、まれに荷物に紛れて輸送されることがあり、日本でも港で発見され、大騒ぎになることがあった。
毒
[編集]
サソリの尾の先には毒針があり、これを使って毒を注入することは一般的によく知られており、猛毒により人が刺されたら死ぬ場合もあるとして恐れられている。神話伝説にも猛毒を持つサソリの話はたびたび出てくる。ギリシア神話では、英雄オーリーオーンを殺してさそり座になったサソリの話が有名である。神話や逸話によりサソリの毒性は誇張された形で認知されている。実際には、ほとんどのサソリの種は大型哺乳類を殺せるほどの猛毒は持っていない。その理由は、サソリは昆虫など小動物を捕食する際に毒を使うことがほとんどであり、大型動物にそれを使うのは防御行動で、本来は大型哺乳類の殺傷を目的としたものではないからである。人間に対して致命的な毒を持つものも存在するが、その数は約1000種類中に僅か25種と少ない。
日本産の種の毒性は低い。日本以外の地域に生息する種でも人命に関わるような毒性を持つものは少ない。しかし、真に危険なものも実際に存在し、サソリによる死者は世界で年間1000人以上とも言われる。また、人家周辺に生息する種もあり、地域によっては被害を受けやすく、南方地域では、靴を履く時に、靴を裏返してサソリがいないかどうか確かめる地域がある。
毒性の弱い種であっても、刺された結果スズメバチの場合と同様アナフィラキシーショックのような症状に陥ることはある。
人命に関わる猛毒をもつ種類はイエローファットテールスコーピオン、ストライプバークスコーピオンなど。この中でも最強の毒をもつのは中東に生息するオブトサソリといわれている。それら強力な毒を持つサソリの多くは、キョクトウサソリ科で占められており、現在これらキョクトウサソリ科のグループは、日本への輸入が原則禁止となっている。密輸事件も起きている。
漢方の生薬学においては、有名なものではトリカブト(附子)などのように、毒性を持つものが独特の効力を発揮するものとして、しばしば生薬に利用されるが、毒性を持つサソリも卒中や神経麻痺・痙攣に効果があるとされる。生薬名を「全蝎」というが、これは生きたままのサソリを食塩水で丸ごと煮てから、全体を乾燥させたものを、生薬として使用されることからこう呼ばれている。
蛍光現象
[編集]
サソリに暗闇でブラックライトを当てると緑色に光る。表皮にあるヒアリン層が蛍光を発するとされるが、これには少なくともβ-カルボリンが関わっている[16]。この蛍光現象はサソリとして一般的であるが、少なくともChaerilidae科の一部の種類は、例外的に蛍光しない[17]。
産まれたてのサソリにはヒアリン層がないが、脱皮を重ねて成長する毎に増え、発光現象が強くなる[18]。液浸標本にしても、周囲にヒアリン層が溶け出して、光るのだという。また、脱皮した後の脱皮殻も光る。
サソリ以外の節足動物、例えばクモ類、昆虫類とヤスデ類の一部も類似の蛍光現象がある[19][20]。
系統と進化
[編集]サソリは鋏角類の中でも系統的位置は多くの説を与えられた分類群である[21]。同じく鋏角類であるがクモガタ類でないウミサソリ類(広翼類)とは形態的に共通点(後体の体節分化と付属肢構成)が多く、これがサソリに近縁であるとの考えはかつてあった(同時にクモガタ類は多系統群となる)[21]。しかしこの見解はの21世紀においては否定的に評価され[21]、両者の共通点は収斂進化の結果であると見なされる[10]。
| |||||||||||||||||||||||||||
| Stomothecata仮説に基づいたサソリの系統的位置。 |
| |||||||||||||||||||||||||||||||||
| 分子系統学に支持されるサソリの系統的位置。 |
サソリは多くのクモガタ類として祖先形質と考えられる特徴をもち、伝統的には最初期に分化したクモガタ類の系統群であると見なされた[3][21]。また、ザトウムシと同様に第1-2脚の顎葉でstomothecaをなし、これに基づいてザトウムシとの類縁関係(Stomothecataを構成する)も提唱された[10][21]。しかし分子系統学[22][23][24][24] と遺伝子重複[25] の解析ではこれらの見解に応じず、むしろ書肺のある他のクモガタ類(四肺類)と姉妹群になる系統関係の方が有力視される(蛛肺類 Arachnopulmonata[23] を構成する)[21]。

-
 Waeringoscorpio westerwaldensis の復元図
Waeringoscorpio westerwaldensis の復元図 -


サソリの系統は水生環境に起源し、基盤的なサソリは書鰓を有する水棲生動物という説は、20世紀においては主流であり[6]、中でも多くの化石サソリを記述した Kjellesvig-Waering 1986 は、全ての基盤的なサソリを水棲であると解釈していた[26]。これによると、クモガタ類の中でサソリと四肺類は別系統で、それぞれ独自に上陸して書肺を収斂進化すると考えられた[21]。しかし21世紀に至って、これらの説は疑問視されつつあり、前述の通りサソリと四肺類の近縁関係は根強く示唆され、解剖学的見解も両者の書肺の連続相同性を支持している[27]。これによると、基盤的なサソリどころか、サソリと四肺類の最も近い共通祖先は既に書肺をもつ陸生クモガタ類であると示される[27][13][21]。加えて、Kjellesvig-Waering 1986 のほとんどの記述も後に単なる過度解釈と疑わしく見受けられ、例えばその文献に水棲で鰓を有すると解釈される基盤的なサソリ Liassoscorpionides、プロスコルピウス(Proscorpius)と Palaeoscorpius は、再検討したところではいずれの化石からも鰓は見当たらなかった[26][6][13]。後者の発見場所は水生環境だが、周りの化石証拠によると海岸線の陸生環境から水中に流れ込んだ可能性が高い[13]。Waeringoscorpio は再検証でもブラシ状の鰓の存在が認められるが、これは原始的な書鰓ではなく、むしろ水生昆虫の鰓のように、陸生から二次的に水生化することによって独自に獲得した派生形質とされる[28]。2020年で既知最古のサソリと解釈されたパリオスコーピオ(Parioscorpio)から現生のサソリと大まかに対応した循環系とされる構造が見られるが、水生であったことは断言できず[1]、後にもサソリどころか、鋏角類ですらない所属亜門不明の節足動物と再記載された[2]。
下位分類
[編集]

.jpg)
_(20812747191).jpg)

以下の分類体系は Dunlop et al. 2020 に基づく(絶滅群は†、科より上位の分類群は太字で示す)[29]。また、和名に関しては日本蠍研究所制定に基づく[30]。
- †(姉妹群)
- エラサソリ亜目 Branchioscorpionina
- †フタイタサソリ下目 Bilobosternina(デボン紀)
- †エラサソリ上科 Branchioscorpionoidea
- †エラサソリ科 Branchioscorpionidae
- †ナガコロシサソリ科 Dolichophonidae
- †エラサソリ上科 Branchioscorpionoidea
- †ソイタサソリ下目 Holosternina(シルル紀 - 三畳紀)
- †トゲサソリ上科 Acanthoscopionoidea(デボン紀)
- †トゲサソリ科 Acanthoscorpionidae
- †セマサソリ科 Stenoscorpionidae(三畳紀)
- †チガイムカシコロシサソリ上科 Allopalaeophonoidea(シルル紀)
- †チガイムカシコロシサソリ科 Allopalaeophonidae
- †ヨアケコロシサソリ上科 Eoctonoidea(石炭紀)
- †チガイウシコロシガタサソリ科 Allobuthiscorpiidae
- †スミサソリ科 Anthracoscorpionidae
- †ウシコロシガタサソリ科 Buthiscorpiidae
- †ヨアケコロシサソリ科 Eoctonidae
- †ガーネットサソリ科 Garnettiidae
- †キョダイサソリ上科 Gigantoscorpionoidea(デボン紀 - 石炭紀)
- †キョダイサソリ科 Gigantoscorpionidae
- †ナミコロシサソリ上科 Mesophonoidea(石炭紀 - 三畳紀)
- †トゲタタカイサソリ科 Centromachidae(石炭紀)
- †科 Gallioscorpionidae(三畳紀)
- †ヌマスミサソリ科 Heloscorpionidae(石炭紀)
- †メゾンサソリ科 Mazoniidae(石炭紀)
- †ナミコロシサソリ科 Mesophonidae(三畳紀)
- †ウィルスサソリ科 Willsiscorpionidae(三畳紀)
- †ムカシサソリ上科 Palaeoscorpioidea(デボン紀 - 三畳紀)
- †ムカシサソリ科 Palaeoscorpionidae
- †カイメンコロシサソリ上科 Spongiophonoidea(デボン紀 - 三畳紀)
- †マエアルクトゥルスサソリ科 Praearcturidae(デボン紀)
- †カイメンコロシサソリ科 Spongiophonidae(三畳紀)
- †トゲサソリ上科 Acanthoscopionoidea(デボン紀)
- †ワレイタサソリ下目 Meristosternina(石炭紀)
- †マルメサソリ上科 Cyclophthalmoidea
- †マルメサソリ科 Cyclophthalmidae
- †コガタクチビルサソリ科 Microlabiidae
- †ムカシウシコロシサソリ上科 Palaeobuthoidea
- †ムカシウシコロシサソリ科 Palaeobuthidae
- †マルメサソリ上科 Cyclophthalmoidea
- †サケイタサソリ下目 Lobosternina(シルル紀 - 石炭紀)
- †オナジウシコロシサソリ上科 Isobuthoidea(石炭紀)
- †ヨアケウシコロシサソリ科 Eobuthidae
- †ヨアケコロシサソリ科 Eoscorpiidae
- †オナジウシコロシサソリ科 Isobuthidae
- †クロノスサソリ科 Kronoscorpionidae
- †アラワレウシコロシサソリ科 Pareobuthidae
- †フクオナジウシコロシサソリ上科 Paraisobuthoidea(石炭紀)
- †フクオナジウシコロシサソリ科 Paraisobuthidae
- †トガリサソリ科 Scoloposcorpionidae
- †ヌマサソリ科 Telmatoscorpionidae
- †サケイタサソリ上科 Loboarchaeoctonoidea(石炭紀)
- †サケイタサソリ科 Loboarchaeoctonidae
- †ウォーターストンサソリ科 Waterstoniidae
- †ムカシコロシサソリ上科 Palaeophonoidea(シルル紀)
- †ムカシコロシサソリ科 Palaeophonidae
- †オナジウシコロシサソリ上科 Isobuthoidea(石炭紀)
- †フタイタサソリ下目 Bilobosternina(デボン紀)
- ゲンセイサソリ亜目 Neoscorpionina
- オレイタサソリ下目 Orthosternina(石炭紀 - 現世)
- ウシコロシサソリ小目 Buthida
- ウシコロシサソリ上科 Buthoidea
- ウシコロシサソリ科 Buthidae
- ヒメミワクサソリ科 Microcharmidae
- ウシコロシサソリ上科 Buthoidea
- チャエリルスサソリ小目 Chaerilida
- チャエリルスサソリ上科 Chaeriloidea
- チャエリルスサソリ科 Chaerilidae
- チャエリルスサソリ上科 Chaeriloidea
- ニセチャクタスサソリ小目 Pseudochactida
- ニセチャクタスサソリ上科 Pseudochactoidea
- ニセチャクタスサソリ科 Pseudochactidae
- ニセチャクタスサソリ上科 Pseudochactoidea
- ドクオサソリ小目 Iurida
- チャクタスサソリ上科 Chactoidea
- ヘブライサソリ科 Akravidae
- ベリサリウスサソリ科 Belisariidae
- チャクタスサソリ科 Chactidae
- マコトサソリ科 Euscorpiidae
- メイシンサソリ科 Superstitionidae
- メナシチャクタスサソリ科 Typhlochactidae
- ルイジサソリ科 Scorpiopidae
- ベジョビスサソリ科 Vaejovidae
- ドクオサソリ上科 Iuroidea
- ムシコロシサソリ科 Caraboctonidae
- アツオサソリ科 Hadruridae
- ドクオサソリ科 Iuridae
- サソリ上科 Scorpionoidea
- ミゾオサソリ科 Bothriuridae
- フタトゲサソリ科 Diplocentridae
- タイラサソリ科 Hemiscorpiidae
- カワリサソリ科 Heteroscorpionidae
- オボソサソリ科 Hormuridae
- ハリオサソリ科 Urodacidae
- シワハサソリ科 Rugodentidae
- サソリ科 Scorpionidae
- チャクタスサソリ上科 Chactoidea
- ウシコロシサソリ小目 Buthida
- オレイタサソリ下目 Orthosternina(石炭紀 - 現世)
- (上位分類未定 エラサソリ亜目内)
- 属 Brontoscorpio(デボン紀)
- 属 Eramoscorpius(シルル紀)
- 属 Gondwanascorpio(デボン紀)
- 属 Gymnoscorpius(石炭紀)
- 属 Hubeiscorpio(デボン紀)
- 属 Liassoscorpionides(ジュラ紀)
- 属 Palaeomachus(石炭紀)
- 属 Permomatveevia(ペルム紀)
- 属 Titanoscorpio(石炭紀)
- 属 Wattisonia(石炭紀)
- (上位分類未定 ゲンセイサソリ亜目内)
- コロシオサソリ亜科 Uroctoninae
主な種類
[編集]日本のサソリ
[編集]以下の2種が知られる。いずれも広域分布種である。
- ヤエヤマサソリ Liocheles australasiae (Fabricius, 1775)
- 2~3cm程度の小型のサソリ。英名は「Dwarf Wood Scorpion」。東南アジア、オセアニア、オーストラリアなどに広く分布。日本では沖縄諸島の一部、宮古諸島のほぼ全域、八重山諸島のほぼ全域に分布。枯れ木の皮の下などに住み、シロアリなどを食べる。雄がほとんど存在せず、単為生殖で殖えるとされている。毒性は弱いとされている。
- マダラサソリ Isometrus maculatus (De Geer, 1778)
- 6cm位の中型のサソリ。英名は「Lesser Brown Scorpion」。世界の熱帯地域に広く分布し、人為的に分布を広げたと思われる。日本では、沖縄の宮古諸島のほぼ全域、八重山諸島のほぼ全域、小笠原諸島の一部に分布し、時折、ホームセンターでの資材の中や、港の倉庫や、積み卸し荷の中で見つかり、パニックになる時もある。
主な飼育されることの多いサソリ
[編集]

.jpg)

- インディアンジャイアントフォレストスコーピオン Gigantometrus swammerdami
- 【和名】スワンメルダムオオハカリサソリ
- サソリ目世界最大種。最大全長は292mmとされている。
- アジアンフォレストスコーピオン Heterometrus spp.
- 【和名】カワリハカリサソリ属の種
- アジアに分布するカワリハカリサソリ属 (Heterometrus) に属する種類の総称。9種類を含んでおり、同定は困難。詳細は以下の通り。コウテイオソレサソリと並んで飼いやすいサソリ。毒性は低いとされている。
-
- チムルマンカワリハカリサソリ Heterometrus cimrmani Kovařík, 2004
- アオハイカワリハカリサソリ Heterometrus glaucus (Thorell, 1876)
- ラオスカワリハカリサソリ Heterometrus laoticus Couzijn, 1981
- ウデナガカワリハカリサソリ Heterometrus longimanus (Herbst, 1800)
- ミノタウロスカワリハカリサソリ Heterometrus minotaurus Plíšková et al., 2016
- ピータースカワリハカリサソリ Heterometrus petersii (Thorell, 1876)
- シレノスカワリハカリサソリ Heterometrus silenus (Simon, 1884)
- トゲハコビカワリハカリサソリ Heterometrus spinifer (Ehrenberg, 1828)
- トレルカワリハカリサソリ Heterometrus thorellii (Pocock, 1897)
- エンペラースコーピオン Pandinus imperator
- 【和名】コウテイオソレサソリ
- 【別名】ダイオウサソリ、オオコガネサソリ
- アフリカ西部に住む大型のサソリ。最大体長は22cmとされている。日本では世界最大種とされることが多いが、実際に最大の記録を持つ種類はスワンメルダムオオハカリサソリ (Gigantometrus swammerdami)。黒色で強靭なハサミをもつ。性質は大人しく、毒性も低いとされており、最も一般的に飼育される。ワシントン条約付属書II類に指定され保護されている。
- レッドクロースコーピオン Pandinoides cavimanus
- 【和名】クボミテジオソレサソリ
- 【別名】レッドクローエンペラースコーピオン
- ダイオウサソリに似る、やや小型の体全体に赤みがかかった種。エンペラースコーピオンと同じくアフリカに生息している。
- サウスアフリカンフラットロックスコーピオン Hadogenes troglodytes troglodytes
- 【和名】ホンアナスミハデスサソリ
- 扁平な体と細い尾が特徴的な大型種。サソリのなかでは長寿。アフリカに分布しており、毒性は低いとされている。
主な危険なサソリ
[編集].jpg)
- ストライプバークスコーピオン Centruroides vittatus
- 【和名】シマジトゲオサソリ
- 4 cmほどの小型種。弱々しくみえるが強力な毒をもつ[31]。フロリダにも近似種のCentruroides gracilisが分布し、いずれも毒性が強い危険な種である。
- デスストーカー Leiurus quinquestriatus
- 【和名】イツスジナメラカオサソリ
- 中近東に生息し、終体第5節が黒いのが特徴。
- 通称「デスストーカー」(Deathstalker)。「オブトサソリ」とされることが多いがこれは「ファットテールスコーピオン」の和訳で、ファットテールスコーピオンとはヒトコロシサソリ属の種類の総称であるので誤り。サソリのなかで最強の毒をもつといわれる[32]。非常に攻撃的で素早い危険なサソリ。
- イエローファットテールスコーピオン Androctonus australis
- 【和名】ミナミヒトコロシサソリ
- 北アフリカに分布する尾の太い中型のサソリ。強い毒を持ち、死亡例もある[33]。近似種のAndroctonus bicolorや、Buthus occitanusと共に危険な種である。ペットとしての人気は高かったが、他のウシコロシサソリ科の種と同様に、後に法律により研究施設等以外での飼育は禁止されている。
- サウスアフリカンジャイアントファットテールスコーピオン Parabuthus transvaalicus
- 【和名】トランスバーグフクウシコロシサソリ
- 【別名】ジャイアントデスストーカー
- ジャイアントデスストーカーとも呼ばれる南アフリカ産の黒い大型サソリ。大型なので注入できる毒の量が多い。近似種のParaduthus lisomaと同じく、毒液を周囲に噴射する[34]。
脚注
[編集]注釈
[編集]出典
[編集]- ^ a b Wendruff, Andrew J.; Babcock, Loren E.; Wirkner, Christian S.; Kluessendorf, Joanne; Mikulic, Donald G. (2020-01-16). “A Silurian ancestral scorpion with fossilised internal anatomy illustrating a pathway to arachnid terrestrialisation” (英語). Scientific Reports 10 (1): 1–6. doi:10.1038/s41598-019-56010-z. ISSN 2045-2322.
- ^ a b Anderson, Evan P.; Schiffbauer, James D.; Jacquet, Sarah M.; Lamsdell, James C.; Kluessendorf, Joanne; Mikulic, Donald G. (2021). “Stranger than a scorpion: a reassessment of Parioscorpio venator, a problematic arthropod from the Llandoverian Waukesha Lagerstätte” (英語). Palaeontology 64 (3): 429–474. doi:10.1111/pala.12534. ISSN 1475-4983.
- ^ a b c d e f g h i j k l m n Lamsdell, James C.; Dunlop, Jason A. (2017). “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3): 395–418. ISSN 1467-8039.
- ^ Lamsdell, James C. (2013-01-01). “Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura”. Zoological Journal of the Linnean Society 167 (1): 1–27. doi:10.1111/j.1096-3642.2012.00874.x. ISSN 0024-4082.
- ^ a b Miether, Sebastian T.; Dunlop, Jason A. (2016-07). “Lateral eye evolution in the arachnids” (英語). Arachnology 17 (2): 103–119. doi:10.13156/arac.2006.17.2.103. ISSN 2050-9928.
- ^ a b c d e f g h Dunlop, Jason A.; Tetlie, O. Erik; Prendini, Lorenzo (2008). “Reinterpretation of the Silurian Scorpion Proscorpius Osborni (whitfield): Integrating Data from Palaeozoic and Recent Scorpions” (英語). Palaeontology 51 (2): 303–320. doi:10.1111/j.1475-4983.2007.00749.x. ISSN 1475-4983.
- ^ Vignoli, Valerio; Prendini, Lorenzo (2009) (英語). Systematic revision of the troglomorphic North American scorpion family Typhlochactidae (Scorpiones, Chactoidea). (Bulletin of the American Museum of Natural History, no. 326). New York : American Museum of Natural History
- ^ “The Scorpion Files - Typhlochactidae”. www.ntnu.no. 2020年12月9日閲覧。
- ^ Shultz, Jeffrey W. (2007-03). “Morphology of the prosomal endoskeleton of Scorpiones (Arachnida) and a new hypothesis for the evolution of cuticular cephalic endoskeletons in arthropods” (英語). Arthropod Structure & Development 36 (1): 77–102. doi:10.1016/j.asd.2006.08.001.
- ^ a b c Dunlop, Jason; Garwood, Russell J. (2014-11-13). “Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders” (英語). PeerJ 2: e641. doi:10.7717/peerj.641. ISSN 2167-8359.
- ^ a b Jeram, Andrew J. (1993/ed). “Scorpions from the Viséan of East Kirkton, West Lothian, Scotland, with a revision of the infraorder Mesoscorpionina” (英語). Earth and Environmental Science Transactions of The Royal Society of Edinburgh 84 (3-4): 283–299. doi:10.1017/S0263593300006106. ISSN 1755-6929.
- ^ a b c d Haug, C.; Wagner, P.; Haug, J.T. (2019-12-31). “The evolutionary history of body organisation in the lineage towards modern scorpions” (英語). Bulletin of Geosciences: 389–408. doi:10.3140/bull.geosci.1750. ISSN 1802-8225.
- ^ a b c d Kühl, Gabriele; Bergmann, Alexandra; Dunlop, Jason; Garwood, Russel J.; Rust, Jes (2012-07). “Redescription and palaeobiology of Palaeoscorpius devonicus Lehmann, 1944 from the Lower Devonian Hunsrück Slate of Germany: PALAEOSCORPIUS DEVONICUS FROM THE HUNSRÜCK SLATE” (英語). Palaeontology 55 (4): 775–787. doi:10.1111/j.1475-4983.2012.01152.x.
- ^ Waddington, Janet; Rudkin, David M.; Dunlop, Jason A. (2015-01-31). “A new mid-Silurian aquatic scorpion—one step closer to land?”. Biology Letters 11 (1): 20140815. doi:10.1098/rsbl.2014.0815. PMC 4321148. PMID 25589484.
- ^ Di, Zhiyong; Edgecombe, Gregory D.; Sharma, Prashant P. (2018-05-21). “Homeosis in a scorpion supports a telopodal origin of pectines and components of the book lungs”. BMC Evolutionary Biology 18 (1): 73. doi:10.1186/s12862-018-1188-z. ISSN 1471-2148. PMC 5963125. PMID 29783957.
- ^ Stachel, Shawn J; Scott A Stockwell and David L Van Vranken (August 1999). “The fluorescence of scorpions and cataractogenesis”. Chemistry & Biology (Cell Press) 6: 531–539. doi:10.1016/S1074-5521(99)80085-4 2008年6月17日閲覧。.
- ^ Lourenço, Wilson R. (2012-12). “Fluorescence in scorpions under UV light; can chaerilids be a possible exception?” (英語). Comptes Rendus Biologies 335 (12): 731–734. doi:10.1016/j.crvi.2012.11.001.
- ^ Hadley, Neil F.; Stanley C. Williams (July 1968). “Surface Activities of Some North American Scorpions in Relation to Feeding”. Ecology (Ecological Society of America) 49 (4): 726–734. doi:10.2307/1935535 2008年6月17日閲覧。.
- ^ “Love Them Or Hate Them, These Ultraviolet Macros Will Change The Way You See Arthropods” (英語). DIY Photography (2015年1月1日). 2019年1月19日閲覧。
- ^ Pearson, Gwen (2013年11月20日). “Luminous Beauty: The Secret World of Fluorescent Arthropods”. Wired. ISSN 1059-1028 2019年1月19日閲覧。
- ^ a b c d e f g h Howard, Richard J.; Edgecombe, Gregory D.; Legg, David A.; Pisani, Davide; Lozano-Fernandez, Jesus (2019-03-01). “Exploring the evolution and terrestrialization of scorpions (Arachnida: Scorpiones) with rocks and clocks” (英語). Organisms Diversity & Evolution 19 (1): 71–86. doi:10.1007/s13127-019-00390-7. ISSN 1618-1077.
- ^ Cunningham, Clifford W.; Martin, Joel W.; Wetzer, Regina; Bernard Ball; Hussey, April; Zwick, Andreas; Shultz, Jeffrey W.; Regier, Jerome C. (2010-02). “Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences” (英語). Nature 463 (7284): 1079–1083. doi:10.1038/nature08742. ISSN 1476-4687.
- ^ a b Giribet, Gonzalo; Wheeler, Ward C.; Hormiga, Gustavo; González, Vanessa L.; Pérez-Porro, Alicia R.; Kaluziak, Stefan T.; Sharma, Prashant P. (2014-11-01). “Phylogenomic Interrogation of Arachnida Reveals Systemic Conflicts in Phylogenetic Signal” (英語). Molecular Biology and Evolution 31 (11): 2963–2984. doi:10.1093/molbev/msu235. ISSN 0737-4038.
- ^ a b Ballesteros, Jesús A.; Sharma, Prashant P. (2019-11-01). “A Critical Appraisal of the Placement of Xiphosura (Chelicerata) with Account of Known Sources of Phylogenetic Error” (英語). Systematic Biology 68 (6): 896–917. doi:10.1093/sysbio/syz011. ISSN 1063-5157.
- ^ Leite, Daniel J.; Baudouin-Gonzalez, Luís; Iwasaki-Yokozawa, Sawa; Lozano-Fernandez, Jesus; Turetzek, Natascha; Akiyama-Oda, Yasuko; Prpic, Nikola-Michael; Pisani, Davide et al. (09 01, 2018). “Homeobox Gene Duplication and Divergence in Arachnids”. Molecular Biology and Evolution 35 (9): 2240–2253. doi:10.1093/molbev/msy125. ISSN 1537-1719. PMC 6107062. PMID 29924328.
- ^ a b Dunlop, Jason A.; Kamenz, Carsten; Scholtz, Gerhard (2007-06-01). “Reinterpreting the morphology of the Jurassic scorpion Liassoscorpionides” (英語). Arthropod Structure & Development 36 (2): 245–252. doi:10.1016/j.asd.2006.09.003. ISSN 1467-8039.
- ^ a b Scholtz, Gerhard; Kamenz, Carsten (2006). “The book lungs of Scorpiones and Tetrapulmonata (Chelicerata, Arachnida): evidence for homology and a single terrestrialisation event of a common arachnid ancestor”. Zoology (Jena, Germany) 109 (1): 2–13. doi:10.1016/j.zool.2005.06.003. ISSN 0944-2006. PMID 16386884.
- ^ Poschmann, Markus; Dunlop, Jason A.; Kamenz, Carsten; Scholtz, Gerhard (2008-12-01). “The Lower Devonian scorpionWaeringoscorpio and the respiratory nature of its filamentous structures, with the description of a new species from the Westerwald area, Germany” (英語). Paläontologische Zeitschrift 82 (4): 418–436. doi:10.1007/BF03184431. OCLC 319031287.
- ^ Dunlop, J. A., Penney, D. & Jekel, D. 2020. A summary list of fossil spiders and their relatives. In World Spider Catalog. Natural History Museum Bern, online at http://wsc.nmbe.ch, version 20.5
- ^ “分類/和名 :: 日本蠍研究所”. japanscorpionlabo.webnode.jp (2023年2月17日). 2023年2月17日閲覧。
- ^ Goetz, David W.; Demain, Jeffrey G. (1995-01-01). “Immediate, late, and delayed skin test responses to Centruroides vittatus scorpion venom” (English). Journal of Allergy and Clinical Immunology 95 (1): 135–137. doi:10.1016/S0091-6749(95)70163-X. ISSN 0091-6749. PMID 7822656.
- ^ Lucian K. Ross (2008年). “Leiurus quinquestriatus (Ehrenberg, 1828)” (PDF). The Scorpion Files. September 17, 2011閲覧。
- ^ “The Scorpion Files - Androctonus australis (Buthidae)”. www.ntnu.no. 2019年2月8日閲覧。
- ^ Nisani, Zia; Hayes, William K. (2015-06-01). “Venom-spraying behavior of the scorpion Parabuthus transvaalicus (Arachnida: Buthidae)”. Behavioural Processes 115: 46–52. doi:10.1016/j.beproc.2015.03.002. ISSN 0376-6357.
参考文献
[編集]- 『毒虫の飼育・繁殖マニュアル』(データハウス)秋山智隆