
節口綱
| 節口綱 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||||||
| 地質時代 | |||||||||||||||
| オルドビス紀(トレマドッグ期) - 現世[1][注釈 1] | |||||||||||||||
| 分類 | |||||||||||||||
| |||||||||||||||
| 学名 | |||||||||||||||
| Merostomata sensu Woodward, 1866 | |||||||||||||||
| 下位分類群[2] | |||||||||||||||
節口綱(せっこうこう)、腿口綱(たいこうこう)、またはメロストマ綱[3](学名: Merostomata)は、鋏角類に属する節足動物の分類群(綱)の1つ。構成種は節口類・腿口類[3](merostome[4])と総称される。カブトガニ類・ウミサソリ類などが含まれ、縁辺部が広げた背甲を特徴とする[5]。
現生では4種のカブトガニ類のみを含むが、化石は300種を超えるほど多く知られる[1]。主に古生代の堆積累層から見つかり、知られる確定的な最古記録は少なくとも約4億8,000万年前のオルドビス紀早期まで遡る[1][注釈 1]。
名称
[編集]学名「Merostomata」はギリシャ語の「meros」(太もも)と「stoma」(口)の合成語。これは本群において、脚の基部が口器(顎基)に特化し、口がその間にあるように見えることに因んでいる[6]。
用法
[編集]1852年、「Merostomata」はジェームズ・デーナ(James Dwight Dana)によって創設され、当時はカブトガニ類のみを含む分類群であった(Merostomata Dana, 1852)。1866年、Henry Woodward はこの分類群にウミサソリ類を追加して以降、節口類(Merostomata sensu Woodward, 1866)は20世紀後期までカブトガニ類とウミサソリ類を含む分類群として広く流用されていた[4]。
しかし20世紀後期以降では、本群の単系統性が一般に否定され、ウミサソリ類はカブトガニ類よりもクモガタ類に近縁という説が主流となっている[5]。このような節口類に対しては、クモガタ類を除いた側系統群とする・節口類を破棄する[7]・カブトガニ類のみを含む Merostomata Dana, 1852 をカブトガニ類 Xiphosura のジュニアシノニム(無効の異名)とする[8]など、様々な意見を与えられる。なお、2010年代以降でもこの分類群の単系統性を支持する研究結果はわずかにあり[5][2]、便宜上の総称として採用し続ける場合もある[9]。
本項目では一般的な用法の節口類、すなわちカブトガニ類やウミサソリ類などを含む Merostomata sensu Woodward, 1866 について扱う。
形態
[編集]

![カブトガニ類の腹面構造[注釈 2]](/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Limulus_ventral_morphology.png)
数cmしか及ばない小型種もあるが、節口類は大型の節足動物を中心とするグループである[10]。現生のカブトガニ類だけでも最大数十cm、絶滅群まで範囲を広げるとウミサソリ類は1m前後の種類が多く、2.5m以上と推測され、発見史上最大級の節足動物として知られるものもある[11]。
数多くの体節は前体と後体という2つの合体節にまとめられ[12]、付属肢(関節肢)は全て体の腹面で覆われる。他の鋏角類(ウミグモ類とクモガタ類)との主な相違点は、縁辺部が出張った背甲・顎基のある脚・可動な蓋板をもつことが挙げられる[5][12]。
前体
[編集]_3_(15113089539).jpg)



前体(prosoma)は先節と第1-6体節の融合でできている合体節で、便宜的に「頭胸部」(cephalothorax)ともいうが、合体節的には頭部そのものに該当する[8]。背面は1枚の背甲(carapace または prosomal dorsal shield[12])に覆われ、これは他の鋏角類よりも発達で、縁辺部が付属肢を覆い被さるほど出張っている[5]。通常、背甲の背面には複眼である側眼(lateral eye)と、単眼である中眼(median eye)をそれぞれ1対もつ。
体節数に応じて、前体には6対の付属肢(関節肢)をもつ。最初の1対は鋏角(chelicera)であり、3節のうち先端2節は鋏をなし、ほとんどの場合は小さく目立たない。次の5対は6節前後に分かれた脚であり、最初のものは触肢(pedipalp)だが、原則としてクモガタ類やウミグモ類ほどには特化しない[12]。これらの脚はカブトガニ類ではほぼ同形だが、ウミサソリ類では番目によって顕著に分化した例が多く見られる。脚の基部は正中線付近で接しており、それぞれ基部の肢節(基節 coxa)は口器の役目をするように顎基(gnathobase)という鋸歯状の内突起をもつものが多い[13]。脚は通常では内肢(endopod)のみをもつ単枝型だが、少なくとも前の4対がはっきりとした外肢(exopod)をもつ二叉型のものもある[12]。
目立たない上唇(labrum)に覆われる口は鋏角と触肢の間に位置するが、カブトガニ類の場合は更に後ろにずらし、口が脚の間にあるように見える。口の直後、いわゆる脚の間には「endostoma」という、クモガタ類の前体の腹板(sternite, sternum)に相同と思われる[12]目立たない外骨格が1枚ある[14][15][16]。
後体
[編集]


-
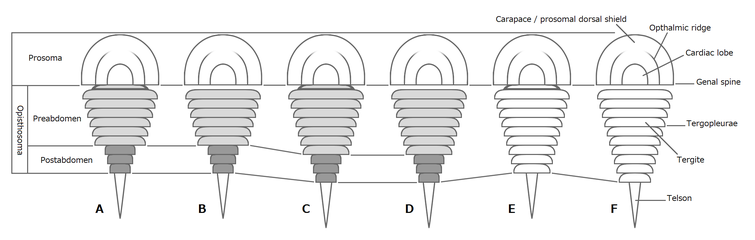 ハラフシカブトガニ類の後体の分化様式
ハラフシカブトガニ類の後体の分化様式

後体(opisthosoma)は第7体節を起点として計9-13節(第15-19体節まで及ぶ)でできた合体節で、便宜的に「腹部」ともいうが、合体節的には胴部(胸部+腹部)に該当する[8]。現生種を含んだカブトガニ亜目は体節がほぼ全て融合しているが、それ以外は体節が明瞭に分節した例が多い[12]。後体は更に体節構造の変化に基づいて、前後で更に幅広い中体(mesosoma, または前腹部 preabdomen)と幅狭い終体(metasoma, または後腹部 postabdomen)として区別できる例もある[8][12]。
後体前7節(第7-13体節)の腹面に附属肢をもつが、最初のもの以外では板状の蓋板(がいばん、operculum)で、その多くが呼吸器である書鰓(しょさい、book gill)をもつ[12]。蓋板の中で最初の1対は生殖孔をもつ生殖口蓋(genital operulum)で、カブトガニ類の場合では1対の単調な生殖孔のみをもつが、ウミサソリ類とカスマタスピス類の場合は特化した生殖肢(genital appendage)をその中央にもつ[12]。
後体第1節(第7体節)は退化的で、背面の外骨格(背板)外見上から観察できない場合が多く、あったとしても幅狭く目立たない[12]。その付属肢は多くの場合では単調な構造体(カブトガニ類の唇様肢 (chilarium)、ウミサソリ類とカスマタスピス類の下層板 (metastoma)[17])であり、前体の一部として機能する傾向が強い[12]。ウェインベルギナは例外的に、この体節に前体のものにほぼ同形な脚をもつとされる[12]。これらの性質に基づいて、この体節を後体ではなく、前体の一部と扱うべきではないかという見解もある(鋏角類#第7体節も参照)[12][9]。
後体の最終体節に続く尾節(telson)は発達で、通常は棘状(尾剣 tail spine)だが、ウミサソリ類ではへら状に特化した例もある[12]。
体節と付属肢比較
[編集]体節 分類
|
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9以降 |
|---|---|---|---|---|---|---|---|---|---|
| オファコルス† | 鋏角 | 脚 (二叉型) |
脚 (二叉型) |
脚 (二叉型) |
脚 (二叉型) |
脚 (分岐あり) |
鰭状の付属肢 | 蓋板 | 蓋板5対 |
| ダイバステリウム† | 鋏角 | 脚 (二叉型) |
脚 (二叉型) |
脚 (二叉型) |
脚 (二叉型) |
脚 | 唇様肢らしい付属肢 | 蓋板(書鰓) | 蓋板(書鰓)5対 |
| ウェインベルギナ† | 鋏角 | 脚 | 脚 | 脚 | 脚 | 脚 | 脚 | 蓋板 | 蓋板5対? |
| カブトガニ亜目 | 鋏角 | 脚 | 脚 | 脚 | 脚 | 脚 (分岐あり) |
唇様肢 | 蓋板 | 蓋板(書鰓)5対 |
| カスマタスピス類† | 鋏角 | 脚 | 脚 | 脚 | 脚 | 脚 | 下層板 | 蓋板(生殖肢) | 蓋板2対 |
| ウミサソリ類† | 鋏角 | 脚 | 脚 | 脚 | 脚 | 脚 | 下層板 | 蓋板(生殖肢) | 蓋板(書鰓)5対 |
| クモガタ類 | 鋏角 | 触肢 | 脚 | 脚 | 脚 | 脚 | -[注釈 3][12] | 蓋板(書肺/生殖肢)/- | 蓋板(書肺)/櫛状板/- |
| ウミグモ類 | 鋏肢 | 触肢 | 担卵肢 | 脚 | 脚 | 脚 | 脚 | - | - |
生態
[編集]節口類は水棲動物であり、現生のカブトガニ類は海棲に限られるが、絶滅群まで範囲を広げると、カブトガニ類とウミサソリ類の両方とも海棲と淡水性の種類が知られている[18][19][20]。多くの種類は捕食者とされる[13]。
一般に脚で水の底を這い回るとされるが、後体の蓋板(カブトガニ類)もしくは特化した遊泳脚(一部のウミサソリ類とカスマタスピス類)で遊泳を行うとされる種類もあり、ウミサソリ類の場合は同時に陸上で活動できたと考えられる種類もある[21]。
分類
[編集]
「鋏角類」という分類群がまだ提唱されない20世紀以前では、節口類は甲殻類と解釈された[22]。後に甲殻類らしからぬ、むしろクモガタ類(クモ・サソリ・ダニなどが属する分類群)に共通な基本体制をもつことが分かり、Heymons 1901 以降ではクモガタ類やウミグモ類とともに鋏角類としてまとめられるようになった[23]。また、本群は複眼・未特化の触肢・発達した後体付属肢と書鰓・二叉型付属肢など、著しく特化したウミグモ類と派生的なクモガタ類には見当たらない、鋏角類の祖先形質(共有原始形質)と思われる特徴が多く整っていたため、鋏角類の基部系統を解明するのに重要視されるグループである[8]。
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Lamsdell (2013)[8] を基に簡略化した鋏角類の内部系統関係。青い枠は節口類に該当する範囲を示す。また、Dekatriata 以外の Prosomapoda の構成群は、側系統群とされる広義のカブトガニ類に該当する(後述参照)[8]。 |
20世紀初期から後期まで、節口類はクモガタ類と共に、真鋏角類(ウミグモ以外の鋏角類)を大きく2つに分ける綱(節口綱・クモガタ綱)とされてきたが、この分類体系は単に真鋏角類を水棲と陸棲に分けるようで、系統関係を反映できるとは限らない問題が指摘される[5]。20世紀後期以降のほとんどの系統解析においても、本群のうちウミサソリ類は(主にクモガタ類に似た生殖器の構造で)カブトガニ類よりもクモガタ類に近縁とされる[24][25][16][5][8][12]。すなわちウミサソリ類とクモガタ類は単系統群(Sclerophorata)をなし、節口類はクモガタ類に至る側系統群とされる[8][12]。
なお、少数であるものの、節口類の単系統性を支持する系統解析もある[5][2]。この場合、節口類は基盤的な鋏角類[5]、もしくはクモガタ類が節口類に対して多系統群とされる[2]。
下位分類
[編集]節口類の構成種は大部分がカブトガニ類とウミサソリ類からなる。それ以外では、かつてカブトガニ類に内包されたカスマタスピス類と、一般に基盤的な真鋏角類とされるオファコルスやダイバステリウムなどが知られている[5][2]。
光楯類はかつて鋏角類と解釈され、20世紀中期まででは節口類(特にカブトガニ類)に分類された[4][8]。しかし再検討により、光楯類はそもそも付属肢が鋏角類らしからぬ(鋏角はなく、触角を含んで頭部付属肢は4-5対しかない)別系統の節足動物だと判明し、20世紀後期以降では三葉虫類と共にArtiopoda類に再分類されるようになった[26]。
カブトガニ類
[編集]
カブトガニ類または剣尾類(カブトガニ目/剣尾目、学名: Xiphosura)[27][28]は、現生は3属4種のみが生存し、化石はオルドビス紀まで遡り[29][30]、80種以上が認められる群である[1]。ウミサソリ類に比べて体は比較的にコンパクトな形をとる。前体の背甲は脚の先端まで覆いかぶさったドーム状に発達し、脚は知られる限り鋏を先端にもつものが多い。6対の蓋板のうち最初の1対は単調な生殖口蓋で、後5対は書鰓をもつ。化石種まで含むと比較的に多様で数cmの小型種が多いが、現生種はいずれも数十cmの大型で、外見・生態ともほぼ共通している[31]。
狭義のカブトガニ類は、後体第1節(第7体節)の背面は外見上から観察できず、その付属肢は小さな唇様肢である[12]。広義のカブトガニ類はハラフシカブトガニ類をも含め、その中でも、後体第1節(第7体節)の付属肢は脚という、他の真鋏角類に類が見られない特徴をもつとされるウェインベルギナがある[8]。カブトガニ亜目の場合、後体第2節(第8体節)以降の体節は全て融合し、「thoracetron」という甲羅をなしている[8][12]。名前の通り、長く伸びていた「尾剣」と呼ばれる剣状の尾節をもつ。
従来は単系統群とされ、カブトガニ亜目(Xiphosurida)とハラフシカブトガニ亜目(ハラフシカブトガニ類/共剣尾類 Synziphosurina)[17]の2群に大きく分かれていた[32]。しかし Lamsdell (2013) の再検討以降では、ハラフシカブトガニ類は系統範囲の雑多な側系統群と見直されるようになった[8]。これにより、従来および広義のカブトガニ類は、ウミサソリ類やクモガタ類など(Dekatriata)を除いた側系統群で、狭義のカブトガニ類は、ハラフシカブトガニ類を除き、カブトガニ亜目といくつかの属のみを含んだ単系統群とされるようになった(カブトガニ類#化石鋏角類との関係性、ハラフシカブトガニ類#分類と進化、および上記の系統図を参照)[8][1][20]。
現生種は全てカブトガニ科(Limulidae)のものであり、東アジアのカブトガニ(Tachypleus tridentatus)、南アジアと南東アジアのミナミカブトガニ(Tachypleus gigas)とマルオカブトガニ(Carcinoscorpius rotundicauda)、および北アメリカのアメリカカブトガニ(Limulus polyphemus)の計4種が知られている。中でアメリカカブトガニは特に研究が進んでいる[31]。基本は底生性だが、蓋板を羽ばたいて逆さまに遊泳することも可能である。雑食性で、二枚貝・巻貝・多毛類・甲殻類などの小動物[33]、および腐肉・海藻を餌とする[34]。





.jpg)

- Xiphosura sensu lato(広義のカブトガニ類、Dekatriataを除いた側系統群)[1]
- Prosomapoda(Dekatriata を除く)
- ウェインベルギナ科 Weinberginidae †
- Planaterga(Dekatriata を除く)
- (科)Bunodidae †
- (科)Pseudoniscidae †
- カブトガニ目 Xiphosura sensu stricto(狭義のカブトガニ類、単系統群)[8][1]
- Prosomapoda(Dekatriata を除く)
ウミサソリ類
[編集]
ウミサソリ類または広翼類(ウミサソリ目/広翼目、学名: Eurypterida)[17]は、およそ250種が認められ[1]、1m前後の大型種を中心とし、既知最大級の節足動物をも含んだ絶滅群である[11]。カブトガニ類に比べて体は細長く、流線型の形をとる。前体の背甲は幅広いが、カブトガニ類ほど極端ではない。鋏角や脚が特化した例が存在し、特にウミサソリ亜目はへら状の遊泳脚に特化した第5脚をもつ。後体は発達で、13節のうち下層板をもつ第1節の背板は観察できないため、背面は12節に見える[12]。6対の蓋板はカブトガニ類のと同じく後5対が書鰓をもつが、腹板と鰓室を構成し、最初の2対は融合して棒状の生殖肢を中央にもつ[12]。後体は更に前後で中体(前腹部)と終体(後腹部)に区別できる場合がある[12]。棘状もしくはへら状の尾節をもつ[12]。
海中および淡水域に生息し、オルドビス紀からペルム紀にかけて生息した。化石から、海中からだんだんと淡水へと生息域を広めていったことが分かる[18]。
古くはカブトガニ類、もしくはクモガタ類のサソリのみに近縁とされてきたが、21世紀以降ではクモガタ類と姉妹群を構成する系統関係の方が広く認められる[24][25][16][8][12]。
-

-

-
 ヒベルトプテルス Hibbertopterus scouleri
ヒベルトプテルス Hibbertopterus scouleri -

-

-

-


.jpg)





- ウミサソリ目 Eurypterida †
- アシナガウミサソリ亜目[17] Stylonurina †
- (上科)Rhenopteroidea †
- (科)Rhenopteridae †
- アシナガウミサソリ上科[17] Stylonuroidea †
- ココモプテルス上科 Kokomopteroidea †
- (上科)Mycteropoidea †
- (上科)Rhenopteroidea †
- ウミサソリ亜目[17] Eurypterina †
- (上科)Onychopterelloidea †
- (科)Onychopterellidae †
- (上科)Moselopteroidea †
- (科)Moselopteridae †
- ヒレオウミサソリ上科[17] Megalograptoidea †
- ウミサソリ上科[17] Eurypteroidea †
- (下目)Diploperculata †
- (上科)Onychopterelloidea †
- アシナガウミサソリ亜目[17] Stylonurina †
カスマタスピス類
[編集]
カスマタスピス類(カスマタスピス目、学名: Chasmataspidida)は、少なくともオルドビス紀からデボン紀にかけて生息し[1]、2019年現在では十数種のみが知られる希少なグループである[35]。ほとんどの種類は3cmを超えない小型で、ウミサソリ類によく似た姿をしているが、後体の体節分化で明確に区別できる[36][12]。鋏角の詳細は不明で、残りの前体付属肢も往々にして化石の保存されていないが、知られるものでは歩脚型で、カスマタスピス(ネジムシ[17])の場合は少なくとも1対が鋏型の先端を有し[36]、ディプロアスピス科の場合は後脚がへら状の遊泳脚に特化した例が確認される[12]。後体は13節からなり、そのうち第1節の背板は消失しないが幅狭い[12]。後体は前4節の前腹部と後9節の後腹部という、他の鋏角類に見当たらない独特な分化様式をもつ[36][12]。カスマタスピスの場合、前腹部の体節はカブトガニ類の後体のように融合が進んでいる[36]。後体腹面は3対の蓋板と、ウミサソリ類に似た下層板と生殖肢をもつ[12]。なお、本群は6対の蓋板をもつことを示唆するカンブリア紀のカスマタスピスらしき生痕化石もある[36]。棘状もしくはへら状の尾節をもつ[12]。このような特徴を兼ね備えるため、かつてカブトガニ類もしくはウミサソリ類と誤同定された種類は少なくない[37][38][39]。
カスマタスピス類の系統位置に対して、古くから様々な仮説(カブトガニ類に含まれる・ウミサソリ類に含まれる・ウミサソリ類に至る側系統群・カブトガニ類とウミサソリ類に対して多系統群など)が提唱された[36][36][35]。しかし2010年代以降では、カスマタスピス類の単系統性、およびウミサソリ類・クモガタ類と共に単系統群(Dekatriata)になる説の方が有力視されつつある(詳細はChasmataspidida#分類を参照)[8][40][41][42]。







その他
[編集]

ここに列挙される種類は、便宜的に広義のカブトガニ類にも含まれる場合もあるが、一般に残りの節口類より基盤的な真鋏角類と見なされる[8][1]。カブトガニ類とウミサソリ類の場合、少なくとも第1-4脚は単枝型で完全に外肢を欠けているが、これらの種類は、発達した歩脚状の外肢を前体第1-4脚に有し、これはハベリアやサンクタカリスのような基盤的な鋏角類から受け継いだ祖先形質と考えられる[12][9]。


脚注
[編集]注釈
[編集]出典
[編集]- ^ a b c d e f g h i j Dunlop, J. A., Penney, D. & Jekel, D. 2020. A summary list of fossil spiders and their relatives. In World Spider Catalog. Natural History Museum Bern, online at http://wsc.nmbe.ch, version 20.5
- ^ a b c d e Ballesteros, Jesús A; Santibáñez-López, Carlos E; Baker, Caitlin M; Benavides, Ligia R; Cunha, Tauana J; Gainett, Guilherme; Ontano, Andrew Z; Setton, Emily V W et al. (2022-02-01). “Comprehensive Species Sampling and Sophisticated Algorithmic Approaches Refute the Monophyly of Arachnida”. Molecular Biology and Evolution 39 (2): msac021. doi:10.1093/molbev/msac021. ISSN 1537-1719.
- ^ a b 小項目事典, ブリタニカ国際大百科事典. “腿口類(たいこうるい)とは - コトバンク”. コトバンク. 2018年11月11日閲覧。
- ^ a b c Eldredge, Niles; Smith, LeGrande (1974) (英語). Revision of the suborder Synziphosurina (Chelicerata, Merostomata) : with remarks on merostome phylogeny. American Museum novitates ; no. 2543.
- ^ a b c d e f g h i j Garwood, Russell J.; Dunlop, Jason (2014-11-13). “Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders” (英語). PeerJ 2: e641. doi:10.7717/peerj.641. ISSN 2167-8359.
- ^ “Virtual museum - Merostomata (Merostomates)”. muzeum.geology.cz. 2018年11月18日閲覧。
- ^ Tudge, Colin (2002) (英語). The Variety of Life: A Survey and a Celebration of All the Creatures that Have Ever Lived. Oxford University Press. ISBN 9780198604266
- ^ a b c d e f g h i j k l m n o p q r Lamsdell, James C. (2013). “Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura” (英語). Zoological Journal of the Linnean Society 167 (1): 1–27. doi:10.1111/j.1096-3642.2012.00874.x. ISSN 0024-4082.
- ^ a b c Aria, Cédric; Caron, Jean-Bernard (2017-12-21). “Mandibulate convergence in an armoured Cambrian stem chelicerate”. BMC Evolutionary Biology 17 (1): 261. doi:10.1186/s12862-017-1088-7. ISSN 1471-2148. PMC 5738823. PMID 29262772.
- ^ Dunlop, Jason A. (2019-01). “Miniaturisation in Chelicerata” (英語). Arthropod Structure & Development 48: 20–34. doi:10.1016/j.asd.2018.10.002.
- ^ a b Braddy, Simon J; Poschmann, Markus; Tetlie, O. Erik (2008-02-23). “Giant claw reveals the largest ever arthropod”. Biology Letters 4 (1): 106–109. doi:10.1098/rsbl.2007.0491. ISSN 1744-9561. PMC 2412931. PMID 18029297.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag A., Dunlop, Jason; C., Lamsdell, James (2017). “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3). ISSN 1467-8039.
- ^ a b Haug, Carolin (2020-08-13). “The evolution of feeding within Euchelicerata: data from the fossil groups Eurypterida and Trigonotarbida illustrate possible evolutionary pathways” (英語). PeerJ 8: e9696. doi:10.7717/peerj.9696. ISSN 2167-8359.
- ^ Selden, Paul A. (1981). “Functional morphology of the prosoma of Baltoeurypterus tetragonophthalmus (Fischer) (Chelicerata: Eurypterida)” (英語). Transactions of the Royal Society of Edinburgh: Earth Sciences 72 (1): 9–48. doi:10.1017/S0263593300003217. ISSN 0263-5933.
- ^ Shultz, J. (2001). GROSS MUSCULAR ANATOMY OF LIMULUS POLYPHEMUS (XIPHOSURA, CHELICERATA) AND ITS BEARING ON EVOLUTION IN THE ARACHNIDA. doi:10.1636/0161-8202(2001)029[0283:GMAOLP]2.0.CO;2.
- ^ a b c Shultz, Jeffrey W. (2007-06-01). “A phylogenetic analysis of the arachnid orders based on morphological characters” (英語). Zoological Journal of the Linnean Society 150 (2): 221–265. doi:10.1111/j.1096-3642.2007.00284.x. ISSN 0024-4082.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y 節足動物の多様性と系統. 石川 良輔 (著, 編集), 馬渡 峻輔 (監修), 岩槻 邦男 (監修). 東京: 裳華房. (2008). ISBN 978-4-7853-5829-7. OCLC 676535371
- ^ a b Tetlie, O. Erik (2007-09-03). “Distribution and dispersal history of Eurypterida (Chelicerata)” (英語). Palaeogeography, Palaeoclimatology, Palaeoecology 252 (3): 557–574. doi:10.1016/j.palaeo.2007.05.011. ISSN 0031-0182.
- ^ Lamsdell, James C. (2016-03). Zhang, Xi-Guang. ed. “Horseshoe crab phylogeny and independent colonizations of fresh water: ecological invasion as a driver for morphological innovation” (英語). Palaeontology 59 (2): 181–194. doi:10.1111/pala.12220.
- ^ a b Bicknell, Russell D. C.; Pates, Stephen (2020). “Pictorial Atlas of Fossil and Extant Horseshoe Crabs, With Focus on Xiphosurida” (English). Frontiers in Earth Science 8. doi:10.3389/feart.2020.00098. ISSN 2296-6463.
- ^ Lamsdell, James C.; McCoy, Victoria E.; Perron-Feller, Opal A.; Hopkins, Melanie J. (2020-11-02). “Air Breathing in an Exceptionally Preserved 340-Million-Year-Old Sea Scorpion” (English). Current Biology 30 (21): 4316–4321.e2. doi:10.1016/j.cub.2020.08.034. ISSN 0960-9822. PMID 32916114.
- ^ Woodward, Henry (1866). A monograph of the British fossil Crustacea, belonging to the order Merostomata.. London,: Printed for the Palæontographical Society,. doi:10.5962/bhl.title.53733
- ^ Heymons, R. (1901年1月1日). “Die Entwicklungsgeschichte der Scolopender” (ドイツ語). www.schweizerbart.de. 2020年11月30日閲覧。
- ^ a b Weygoldt, P.; Paulus, H. F. (1979). “Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata1 II. Cladogramme und die Entfaltung der Chelicerata” (英語). Journal of Zoological Systematics and Evolutionary Research 17 (3): 177–200. doi:10.1111/j.1439-0469.1979.tb00699.x. ISSN 1439-0469.
- ^ a b Shultz, Jeffrey W. (1990). “Evolutionary Morphology and Phylogeny of Araghnida” (英語). Cladistics 6 (1): 1–38. doi:10.1111/j.1096-0031.1990.tb00523.x. ISSN 1096-0031.
- ^ Ortega-Hernández, Javier; Legg, David A.; Braddy, Simon J. (2012-08-15). “The phylogeny of aglaspidid arthropods and the internal relationships within Artiopoda” (英語). Cladistics 29 (1): 15–45. doi:10.1111/j.1096-0031.2012.00413.x. ISSN 0748-3007.
- ^ 小項目事典, ブリタニカ国際大百科事典. “カブトガニ類(カブトガニるい)とは - コトバンク”. コトバンク. 2018年11月15日閲覧。
- ^ 第2版,日本大百科全書(ニッポニカ), ブリタニカ国際大百科事典 小項目事典,デジタル大辞泉,世界大百科事典. “剣尾類(けんびるい)とは - コトバンク”. コトバンク. 2018年11月15日閲覧。
- ^ The oldest horseshoe crab: A new xiphosurid from Late Ordovician Konservat-Lagerstatten deposits, Manitoba, Canada
- ^ “(PDF) A new Ordovician arthropod from the Winneshiek Lagerstätte of Iowa (USA) reveals the ground plan of eurypterids and chasmataspidids” (英語). ResearchGate. 2018年11月18日閲覧。
- ^ a b Dolejš, Petr; Vaňousová, Kateřina (2015-06-01). A collection of horseshoe crabs (Chelicerata: Xiphosura) in the National Museum, Prague (Czech Republic) and a review of their immunological importance. 49. pp. 1–9. doi:10.5431/aramit4901.
- ^ ondrej.zicha(at)gmail.com, Ondrej Zicha;. “BioLib: Biological library” (チェコ語). www.biolib.cz. 2018年11月10日閲覧。
- ^ “(PDF) Diet and food preferences of the adult horseshoe crab Limulus polyphemus in Delaware Bay, New Jersey, USA” (英語). ResearchGate. 2018年11月15日閲覧。
- ^ “Horseshoe Crabs, Limulus polyphemus”. MarineBio.org. 2018年11月15日閲覧。
- ^ a b Lamsdell, James C.; Gunderson, Gerald O.; Meyer, Ronald C. (2019-01-08). “A common arthropod from the Late Ordovician Big Hill Lagerstätte (Michigan) reveals an unexpected ecological diversity within Chasmataspidida”. BMC Evolutionary Biology 19 (1): 8. doi:10.1186/s12862-018-1329-4. ISSN 1471-2148.
- ^ a b c d e f g Jason A. Dunlop, Lyall I. Anderson & Simon J. Braddy (2004). “A redescription of Chasmataspis laurencii Caster & Brooks (Chelicerata: Chasmataspidida) from the Middle Ordovician of Tennessee, USA, with remarks on chasmataspid phylogeny” (PDF). Transactions of the Royal Society of Edinburgh: Earth Sciences 94 (4): 207–225. doi:10.1017/S0263593300000626.
- ^ Dunlop, Jason A. (2002) (英語). Arthropods from the Lower Devonian Severnaya Zemlya Formation of October Revolution Island (Russia).
- ^ Marshall, David J.; Lamsdell, James C.; Shpinev, Evgeniy; Braddy, Simon J. (2014). “A diverse chasmataspidid (Arthropoda: Chelicerata) fauna from the Early Devonian (Lochkovian) of Siberia” (英語). Palaeontology 57 (3): 631–655. doi:10.1111/pala.12080. ISSN 1475-4983.
- ^ Lamsdell, James C. (2019-11-04). “A chasmataspidid affinity for the putative xiphosuran Kiaeria Størmer, 1934” (英語). PalZ. doi:10.1007/s12542-019-00493-8. ISSN 1867-6812.
- ^ Selden, Paul A.; Lamsdell, James C.; Qi, Liu (2015). “An unusual euchelicerate linking horseshoe crabs and eurypterids, from the Lower Devonian (Lochkovian) of Yunnan, China” (英語). Zoologica Scripta 44 (6): 645–652. doi:10.1111/zsc.12124. ISSN 1463-6409.
- ^ Lamsdell, James C.; Briggs, Derek E. G.; Liu, Huaibao P.; Witzke, Brian J.; McKay, Robert M. (2015-10). “A new Ordovician arthropod from the Winneshiek Lagerstätte of Iowa (USA) reveals the ground plan of eurypterids and chasmataspidids” (英語). The Science of Nature 102 (9-10): 63. doi:10.1007/s00114-015-1312-5. ISSN 0028-1042.
- ^ Lamsdell, James C. (2016-03). Zhang, Xi-Guang. ed. “Horseshoe crab phylogeny and independent colonizations of fresh water: ecological invasion as a driver for morphological innovation” (英語). Palaeontology 59 (2): 181–194. doi:10.1111/pala.12220.