
「マツ材線虫病」の版間の差分
INAKAvillage211 (会話 | 投稿記録) m →外部リンク: リンク切れのものを整理 |
m 外部リンクの修正 (laws.e-gov.go.jp) (Botによる編集) |
||
| (29人の利用者による、間の80版が非表示) | |||
| 1行目: | 1行目: | ||
'''マツ材線虫病'''(英名:'''pine wilt disease''')とは、[[マツ属]]([[学名]]:''Pinus'')樹木に発生する感染症である。病原体は北米原産で[[日本]]を含むアジアやヨーロッパのマツ類に枯死を伴う激害をもたらしている。日本 |
'''マツ材線虫病'''(マツざいせんちゅうびょう、英名:'''pine wilt disease''')とは、[[マツ属]]([[学名]]:''Pinus'')を中心とした[[マツ科]]樹木に発生する線虫の[[感染症]]である。病原体は北米原産で[[日本]]を含むアジアやヨーロッパのマツ類に枯死を伴う激害をもたらしている。日本における病気の汚染地域は徐々に拡大しており、2010年以降、本州以南の46都府県全てで確認されている。関係者の間では'''マツ枯れ'''と呼ばれることが多い。本項でも「マツ枯れ」を用いる。行政用語としては'''松くい虫'''が用いられる。 |
||
世界三大樹木病害([[ニレ立枯病]]、[[クリ胴枯病]]、[[五葉マツ類発疹さび病]])に本病を加えて世界四大樹木病害と呼ぶことがある。また、外見上類似した病害として[[ナラ枯れ]](ブナ科樹木萎凋病)がある。 |
世界三大樹木病害([[ニレ立枯病]]、[[クリ胴枯病]]、[[五葉マツ類発疹さび病]])に本病を加えて世界四大樹木病害と呼ぶことがある。また、外見上類似した病害として[[ナラ枯れ]](ブナ科樹木萎凋病)がある。 |
||
== 症状 == |
== 症状と診断 == |
||
典型的な症状としては盛夏から秋にかけてそれまで正常であったマツの針葉が急速に色あせて最終的には褐変する。針葉の褐変は症状の最終段階であり、それに先だって外見は正常なまま樹脂(いわゆる[[松脂]])の滲出が減少する。健康なマツは幹に傷を付けると大量の樹脂を傷口に分泌するが、この病気を発病したマツは樹脂の量が減少し全く出さないことも多い。このため、早期の診断には幹にピンを刺したりポンチで穿孔したりして樹脂滲出異常の有無を調べる。 |
典型的な症状としては盛夏から秋にかけてそれまで正常であったマツの針葉が急速に色あせて最終的には褐変する。針葉の褐変は症状の最終段階であり、それに先だって外見は正常なまま樹脂(いわゆる[[松脂]])の滲出が減少する。健康なマツは幹に傷を付けると大量の樹脂を傷口に分泌するが、この病気を発病したマツは樹脂の量が著しく減少し全く出さないことも多い。このため、早期の診断には幹にピンを刺したりポンチで穿孔したりして樹脂滲出異常の有無を調べる<ref>日塔正俊, 小田久五, 加藤幸雄 (1967)、「[https://cir.nii.ac.jp/crid/1572824502059207040 522マツ類の穿孔虫に関する研究: 加害対象木の判定と季節的推移について(第78回日本林学会大会)]」 『日本林学会誌』 1967年 49号 3号 p.126-126</ref><ref>小田久五 (1967) 「松くい虫の加害対象木とその判定法について」 『森林防疫ニュース』16:263-266, {{naid|10009947486}}</ref>。この簡易判定方法は発見者の小田久五に因み「小田式判定法」などと呼ばれる。また、発病した個体の幹には多数の穴が見られることが多い。これはこの病気に限ったことではないが、マツが弱ってくると[[キクイムシ]]や[[カミキリムシ]]が集まってくるためである。 |
||
また、発病した個体の幹には多数の穴が見られることが多い。これはこの病気に限ったことではないが、マツが弱ってくると[[キクイムシ]]や[[カミキリムシ]]が集まってくるためである。 |
|||
条件によっては典型的な経過とならず、樹脂滲出が止まっても外見が正常なまま翌年まで生存することがある。冷涼な地方ではこのような経過をたどる個体が温暖な地方より多い。それらの個体は翌年の春から初夏に枯死して「年越し枯れ」と呼ばれたり、さらに遅れて通常のマツ枯れシーズンに至って枯れて「潜在感染木」と呼ばれることもある。 |
条件によっては典型的な経過とならず、樹脂滲出が止まっても外見が正常なまま翌年まで生存することがある。冷涼な地方ではこのような経過をたどる個体が温暖な地方より多い。それらの個体は翌年の春から初夏に枯死して「年越し枯れ」と呼ばれたり、さらに遅れて通常のマツ枯れシーズンに至って枯れて「潜在感染木」と呼ばれることもある。 |
||
多数の個体が同時に発病するとさながら[[紅葉]]のような様相を呈する。しかし本物の紅葉とは異なり翌年の芽吹きはなく、林分内のマツは数年のうちに次々と枯損して大多数の個体が失われる。枯損木はよく風雨にさらされ[[樹皮]]を失って「白骨化」し、徐々に朽ちてゆく。 |
|||
<gallery> |
<gallery> |
||
File:Big Austrian pine with pine wilt.jpg|枯死したヨーロッパクロマツ''P. nigra'' |
File:Big Austrian pine with pine wilt.jpg|枯死したヨーロッパクロマツ''P. nigra'' |
||
File:Bursaphelenchus xylophilus in Bonin Islands.jpg|枯死したリュウキュウマツ(''P. luchuensis'') |
File:Bursaphelenchus xylophilus in Bonin Islands.jpg|枯死したリュウキュウマツ(''P. luchuensis'') |
||
File:Austrian pine with wilt large.jpg|葉が褐色に変色 |
File:Austrian pine with wilt large.jpg|葉が褐色に変色したヨーロッパクロマツ(''P. nigra'') |
||
</gallery> |
</gallery> |
||
== 原因 == |
== 原因 == |
||
[[マツノザイセンチュウ]] ''Bursaphelenchus xylophilus'' (Steiner & Buhrer) Nickleと呼ばれる[[線形動物|線虫]]の一種の感染による。以下、特に断りのない限り線虫と略す。かつてはマツ枯れの原因は昆虫ではないかという説が長くあり、「松 |
[[マツノザイセンチュウ]] ''Bursaphelenchus xylophilus'' (Steiner & Buhrer) Nickleと呼ばれる[[線形動物|線虫]]の一種の感染による。以下、特に断りのない限り線虫と略す。かつてはマツ枯れの原因は昆虫ではないかという説が長くあり、「松くい虫」などの病名にも名残が見られる。この説では「松くい虫」として何種類かの[[キクイムシ]]や[[ゾウムシ]]、[[カミキリムシ]]などが挙げられており防除方法の研究が行われていた<ref>笹川満廣・脇田棘三(1959)アカマツ穿孔ゾウムシ類の加害に関する一観察. 京都府立大學學術報告 農學11:71-74.</ref>。確かに枯死木には多数の昆虫が見られるが、これらはマツが弱ってから侵入したのであって因果が逆であることが分かっている。また、枯死木を伐採するとその切り口が青色に変色していることが多いことから、これを引き起こす青変菌がマツ枯れの原因ではないかという説もあった。1969年、林業試験場(当時、現:[[森林総合研究所]])九州支所の職員だった清原友也と徳重陽山の2人によって[[九州]]のマツ枯れ枯死木から日本では見慣れない線虫が非常に高い確率で発見される現象が報告され<ref>徳重陽山, 清原友也(1969)[https://cir.nii.ac.jp/crid/1390001206355304320 マツ枯死木中に生息する線虫''Bursaphelenchus'' sp.] 『日本林學會誌』 51(7), 193-195.</ref>、さらにその後この線虫を接種したマツが枯死したことから線虫がマツを枯死させる原因であることが判明<ref name="マツ生立木の対する線虫接種試験">清原友也, 徳重陽山(1971)「[https://doi.org/10.11519/jjfs1953.53.7_210 マツ生立木に対する線虫''Bursaphelenchus'' sp.の接種試験]」 『日本林学会誌』 1971年 53巻 7号 p.210-218, {{doi|10.11519/jjfs1953.53.7_210}}</ref>。この線虫は線虫学者の真宮靖治らによって新種''Bursaphelenchus lignicolous''として記載され<ref>Mamiya, Y., and Kiyohara, T., (1972) Description of ''Bursaphelenchus lignicolus'' n. sp. (Nematoda: Aphelenchoidae) from pine wood and histopathlogy of nematode infected trees. Nematologica18(1) :120-124. </ref>、和名はマツノザイセンチュウとされた<ref>真宮靖治, 清原友也, 徳重陽山(1971)「[https://cir.nii.ac.jp/crid/1571980076757607040 マツの材組織中にみられる''Bursaphelenchus''属線虫の1新種, マツノザイセンチュウ(仮称)]」 『日本応用動物昆虫学会大会講演要旨』 (15), 28</ref>。 |
||
線虫の感染からマツの枯死に至るメカニズムは完全には解明されていないが、最終的には樹体内部の管類の閉塞、特に[[仮道管]]の閉塞が広範囲で起こり水を吸い上げられなくなって枯死することが分かっている。抵抗性種では線虫の移動が抑制され管類の閉塞が部分的であり、致命的な全身の通水障害を起こさないために枯死には至らないという。共に抵抗性だが分類学的には遠縁の[[ストローブマツ]](''Pinus strobus'')と[[テーダマツ]](''P. taeda'')を観察し比較した結果では、抵抗性種であっても種類によってその防御機構が異なっていることを示唆するような報告がある<ref>山田利博, 伊藤進一郎 (1993)、「[https://doi.org/10.3186/jjphytopath.59.666 材線虫病抵抗性マツ,テーダマツおよびストローブマツ,のマツノザイセンチュウ感染に対する化学的防御反応]」 『日本植物病理学会報』 1993年 59巻 6号 p.666-672, {{doi|10.3186/jjphytopath.59.666}}, 日本植物病理学会</ref>。 |
|||
近年、この閉塞の原因は線虫や細胞による物理的な管の詰まりではなく、[[キャビテーション]]という現象によって細い管内に気泡が発生することにより管内部に空洞が形成され、この空洞が栓の役割を果たして樹液の流れを妨げること(ガスエンボリズム=気体塞栓症)が継続的に発生していることが明らかになってきた<ref>黒田慶子, 山田利博, 峯尾一彦, 田村弘忠、「[https://doi.org/10.3186/jjphytopath.54.606 マツ材線虫病の病徴進展におけるキャビテーションの影響]」 『日本植物病理学会報』 1988年 54巻 5号 p.606-615, {{doi|10.3186/jjphytopath.54.606}}, 日本植物病理学会</ref><ref>福田健二, 寳月岱造, 鈴木和夫、「[https://agriknowledge.affrc.go.jp/RN/2010510343 病原性の異なるマツノザイセンチュウおよびニセマツノザイセンチユウ接種したクロマツ木部の解剖学的変化および通導阻害]」『日本林學會誌』 1992年 74巻 4号 p.289-299, 日本林學會</ref>。キャビテーションによる仮道管の閉塞自体は健全個体でも乾燥時等に見られるが一時的であれば可逆的である。なぜ継続的なキャビテーションが発生してしまうのかは分かっていない。線虫もしくは線虫に抵抗したマツが作り出す毒素説、菌類などの第三者が関わる説、樹液が変性し[[表面張力]]を失う説などがある。 |
|||
キャビテーション・エンボリズムの診断方法として着色溶液をマツに吸わせてから切断して断面を観察するという古典的方法の他にも、水が途切れる際に発生する音の一種、[[アコースティック・エミッション]](Acoustic Emission, AE)を観測するという非破壊的方法が提案・実用化されており<ref>池田武文 (1994)「[https://doi.org/10.11519/jjfs1953.76.4_364 樹木に発生するキャビテーションのアコースティック・エミッション法による検出]」 『日本林学会誌』 1994年 76巻 4号 p.364-366, {{doi|10.11519/jjfs1953.76.4_364}}, 日本森林学会</ref>、蒸散速度や光合成速度の低下を観察するという従来からの非破壊的方法に加えてこのような面からも樹木の水分異常を観測できるようになってきている。 |
|||
線虫は病原性を持ちマツを枯らすことが出来るが、他のマツへの移動手段を持たない。これを助けるのがヒゲナガカミキリ属(''Monochamus'')のカミキリムシである。線虫は蛹室内にいるカミキリムシ新成虫の気門に侵入し、脱出したカミキリムシと共に他のマツへと移る。 |
|||
線虫の感染からマツの枯死に至るメカニズムは完全には解明されていないが、最終的には樹体内部の管類の閉塞、特に[[仮道管]]の閉塞が広範囲で起こり水を吸い上げられなくなって枯死することが分かっている。抵抗性種では線虫の移動が抑制され管類の閉塞が部分的であり、致命的な全身の通水障害を起こさないために枯死には至らないという。共に抵抗性だが分類学的には遠縁の[[ストローブマツ]](''Pinus strobus'')と[[テーダマツ]](''P. taeda'')を観察し比較した結果では、抵抗性種であっても種類によってその防御機構が異なっていることを示唆するような報告がある<ref>[http://ci.nii.ac.jp/naid/110002736651 T. Yamada and S. Ito (1993)Chemical Defense Responses of Wilt-Resistant Pine Species,''Pinus strobus'' and ''P.taeda'',against ''Bursaphelenchus xylophilus'' Infection. 日本植物病理學會報 59(6), 666-672.]</ref>。 |
|||
=== 病原線虫 ''Bursaphelenchus xylophilus'' === |
|||
近年、この閉塞の原因は線虫や細胞による物理的な管の詰まりではなく、[[キャビテーション]]と言って細い管内に気泡が発生することにより管内部に空洞が形成され、樹液の流れを妨げること(エンボリズム)が継続的に発生していることが明らかになってきた<ref>[http://ci.nii.ac.jp/naid/110002748419 黒田慶子・山田利博・峯尾一彦・田村弘忠.(1989)マツ材線虫病の病徴進展におけるキャビテーションの影響. 日本植物病理学会報54(5),606-615.]</ref>。キャビテーションによる仮道管の閉塞自体は健全個体でも見られるが一時的である。なぜ継続的なキャビテーションが発生するのかは分かっていない。線虫もしくは線虫に抵抗したマツが作り出す毒素説、、菌類などの第三者が関わる説、樹液が変性し[[表面張力]]を失う説などがある。 |
|||
この線虫が初めて見つかったのはアメリカ・[[ルイジアナ州]]で、現地原産の[[ダイオウマツ]](''Pinus palustris'')木材中から発見され、当初は''Aphelenchoides xylophilus''として記載された<ref>Steiner G., Burhrer E. M., (1934) ''Aphelenchoides xylophilus'', n. sp., a nematode associated with blue-stain and other fungi in timber. Journal of Agriculture Research 48944951.</ref>が、その後''Bursaphelenchus''属に移された。日本で見つかりマツへの病原性を示す線虫は前述のように''Bursaphelenchus lignicolous''として新種記載されたが、これも後に移動され同一種扱いとなっている。 |
|||
''B. xylophilus''は体長1mmほどの三日月形で雌雄の区別がある。餌はマツの[[柔組織|柔細胞]]と[[菌類]]であり、[[セルロース]]分解酵素(セルラーゼ)を分泌してマツの細胞を破壊・摂取する。実験室内では菌類だけを餌に培養することが可能である。実験室内での培養温度は餌にする菌類の種類にもよるが15℃以上で増殖、25℃程度で活発に増殖するという<ref>堂園安生・清原友也 (1971) 菌糸培養法におけるマツノザイセンチュウの増殖温度. 日本森林学会九州支部研究論文 25: 160-161. </ref>。ただし、何世代にもわたって実験室内で培養し続けると病原性と増殖力が著しく低下してしまうという報告がある<ref name="清原 (1976)">清原友也 (1976)「[https://doi.org/10.14855/jjn1972.6.56 マツノザイセンチュウの継代培養による病原性の低下]」 『日本線虫学会誌』 1976年 6巻 p.56-59, {{doi|10.14855/jjn1972.6.56}}, 日本線虫学会</ref>。野外においても病原性の異なる系統が共存しているという<ref>Kiyohara, T., and Bolla, R., I., (1990) "[https://academic.oup.com/forestscience/article-abstract/36/4/1061/4642706 Pathogenic Variability Among Populations of the Pinewood Nematode, ''Bursaphelenchus Xylophilus.]''", Forest Science 36(4): 1061-1076.<!--参照先のdoiには誤りがあり飛べないので普通の url を記述--></ref>。病原性の違いの一つにセルロース分解酵素の強弱の差があるとされる<ref>小島克己, 上條厚, 益守眞也, 佐々木惠彦 (1994)、「[https://doi.org/10.11519/jjfs1953.76.3_258 病原力の異なるマツノザイセンチュウの分泌するセルラーゼの性質]」<!--誤植と考えられるが書誌情報頁は"牲質"と記載--> 『日本森林学会誌』 1994年 76巻 3号 p.258-262, {{doi|10.11519/jjfs1953.76.3_2587}}</ref>。 |
|||
線虫は病原性を持ちマツを枯らすことが出来るが、他のマツへの移動手段を持たない。これを助けるのがヒゲナガカミキリ属(''Mochamus'')のカミキリムシである。線虫はカミキリムシが蛹の時期のカミキリムシの気門に侵入し、羽化したカミキリムシと共に他のマツへと移る。 |
|||
カミキリムシ(以下カミキリ)の蛹の気門に入った線虫はカミキリ成虫が羽化するまで待つ。カミキリが羽化し、成虫が餌として齧る健康なマツの若枝から健康なマツの樹体内に侵入する<ref name="森本桂・岩崎厚(1972)">森本桂, 岩崎厚、「[https://doi.org/10.11519/jjfs1953.54.6_177 マツノザイセンチュウ伝播者としてのマツノマダラカミキリの役割]」 『日本林學會誌』 1972年 54巻 6号 p.177-183, {{doi|10.11519/jjfs1953.54.6_177}}, 日本森林学会</ref>。この時期は後述のカミキリの生活環にもよるが、一般に5―8月の初夏~盛夏の時期である。マツ体内では樹脂道を使い移動<ref>真宮靖治(1980)、「[https://doi.org/10.11519/jjfs1953.62.5_176 マツノザイセンチュウの接種による当年生アカマツ苗の発病とその病態解剖]」 『日本林学会誌』 1980年 62巻 5号 p.176-183, {{doi|10.11519/jjfs1953.62.5_176}}, 日本森林学会</ref>、交尾・産卵を繰り返しながら増殖する。このとき抵抗性の種では線虫の移動は部分的であるが、感受性の種は全身に移動し分布するようになる。また、線虫の増殖は感受性種であっても最初は穏やか、マツが目に見えて弱り始めてから爆発的に増えると言う。マツは線虫の侵入(感染)から2週間程度で樹脂の分泌が減少を始め、2-3カ月程度で急激な葉の変色と枯死に至る。弱ったマツはカミキリの格好の産卵場所であり、カミキリが産卵に来る。線虫は枯死木の中で青変菌などを食べながら増殖し<ref name="マツノザイセンチュウの生活環に関連する糸状菌(1)">小林享夫, 佐々木克彦, 真宮靖治(1974)、「[https://doi.org/10.11519/jjfs1953.56.4_136 マツノザイセンチュウの生活環に関連する糸状菌 (I)]」 『日本林學會誌』 1974年 56巻 4号 p.136-145, {{doi|10.11519/jjfs1953.56.4_136}}, 日本森林学会</ref><ref name="マツノザイセンチュウの生活環に関連する糸状菌(2)">小林享夫・佐々木克彦・真宮靖治(1975)、「[https://doi.org/10.11519/jjfs1953.57.6_184 マツノザイセンチュウの生活環に関連する糸状菌 (II)]」 『日本林學會誌』 1975年 57巻 6号 p.184-193, {{doi|10.11519/jjfs1953.57.6_184}}, 日本森林学会</ref>カミキリに乗り移る機会を窺う。年が明けカミキリが蛹から羽化して成虫になる際に線虫はカミキリ気門に潜り込みカミキリと共に新しいマツへと移る。 |
|||
=== 線虫 === |
|||
元々は北米原産種で1930年代にアメリカ産の[[ダイオウマツ]](''Pinus palustris'')から発見された。体長1mmほどの三日月形で雌雄の区別がある。餌はマツの[[柔組織|柔細胞]]と[[菌類]]であり、実験室内では菌類を餌に培養することが可能である。カミキリムシ(以下カミキリ)の蛹の気門に入った線虫はカミキリ成虫が羽化するまで待つ。カミキリが羽化し、成虫が餌として齧る健康なマツの若枝から健康なマツの樹体内に侵入する。この時期は後述のカミキリの生活環にもよるが、一般に5―8月の初夏~盛夏の時期である。マツ体内では樹脂道を使い移動、交尾・産卵を繰り返しながら増殖する。このとき抵抗性の種では線虫の移動は部分的であるが、感受性の種は全身に移動し分布するようになる。また、線虫の増殖は感受性種であっても最初は穏やか、マツが目に見えて弱り始めてから爆発的に増えると言う。マツは線虫の侵入(感染)から2週間程度で樹脂の分泌が減少を始め、2-3カ月程度で急激な葉の変色と枯死に至る。弱ったマツはカミキリの格好の産卵場所であり、カミキリが産卵に来る。線虫は枯死木の中で青変菌などを食べながら増殖しカミキリに乗り移る機会を伺う。年が明け産みつけられた卵が成長し蛹になると線虫は蛹の気門に潜り込み新しいマツへの感染を繰り返す。 |
|||
<gallery> |
<gallery> |
||
File:Bursaphelenchus xylophilus male tail.jpg|雄線虫 |
File:Bursaphelenchus xylophilus male tail.jpg|雄線虫 |
||
File:Bursaphelenchus xylophilus spicule.jpg|雄線虫と突起状の生殖器 |
File:Bursaphelenchus xylophilus spicule.jpg|雄線虫と突起状の生殖器 |
||
File:Berlese.svg|線虫を採取するベールマン装置模式図 |
|||
</gallery> |
</gallery> |
||
==== 類縁の在来線虫 ==== |
|||
マツノザイセンチュウは北米原産の外来種とされるが、日本には在来種として同属のニセマツノザイセンチュウ ''B. mucronatus'' Mamiya et Enda が分布している。本種はほとんど病原性を持たず自らマツに病気を起こすことはできないため、すでに何らかの理由で衰弱していたマツを生息場所にしている<ref>石橋信義 編著. (2003) 線虫の生物学. 東京大学出版会. 東京.</ref>。マツノマダラカミキリも同様で、衰弱して樹脂などによる防御力の弱まったマツに産卵するのが本来の生活様式であった。ニセマツノザイセンチュウもマツノマダラカミキリを媒介者とするが、マツノザイセンチュウとは異なりカミキリを離脱するのは後食時ではなく主に産卵時である。つまり、健全なマツを発病させて媒介者であるカミキリを呼ぶ代わりに、衰弱したマツに媒介者と同時に侵入していると考えられている。ニセマツノザイセンチュウとマツノマダラカミキリは、マツ林に時折生ずる被圧木や折損枝、風倒木などの衰弱木や新鮮枯死材を利用して、細々と生活していた生物であると考えられている。 |
|||
==== ニセマツノザイセンチュウ ''Bursaphelenchus mucronatus'' ==== |
|||
=== カミキリムシ === |
|||
マツノザイセンチュウは北米原産の外来種とされるが、日本には在来種として同属のニセマツノザイセンチュウ ''B. mucronatus'' Mamiya et Enda が分布している。本種と比べると弱い病原性を持つ<ref>神崎菜摘, 二井一禎、「[https://www.jstage.jst.go.jp/article/ajsaez/47/0/47_0_157_1/_article/-char/ja ニセマツノザイセンチュウのアカマツに対する病原性]」 『日本応用動物昆虫学会大会講演要旨』 2003年 47巻 日本応用動物昆虫学会大会講演要旨, p.157, 日本応用動物昆虫学会</ref>が自らマツに病気を起こすことはできないため、すでに何らかの理由で衰弱していたマツを生息場所にしている<ref>石橋信義 編著. (2003) 線虫の生物学. 東京大学出版会. 東京.</ref>。マツノマダラカミキリも同様で、衰弱して樹脂などによる防御力の弱まったマツに産卵するのが本来の生活様式であった。ニセマツノザイセンチュウもマツノマダラカミキリを媒介者とするが、マツノザイセンチュウとは異なりカミキリを離脱するのは後食時ではなく主に産卵時である。つまり、健全なマツを発病させて媒介者であるカミキリを呼ぶ代わりに、衰弱したマツに媒介者と同時に侵入していると考えられている。ニセマツノザイセンチュウとマツノマダラカミキリは、マツ林に時折生ずる被圧木や折損枝、風倒木などの衰弱木や新鮮枯死材を利用して、細々と生活していた生物であると考えられている。 |
|||
この線虫を媒介するカミキリムシは[[ヒゲナガカミキリ属]](''Mochamus'')に属する。日本ではマツノマダラカミキリ(''Mochamus alternatus'')やカラフトヒゲナガカミキリ(''M. saltuaris'')が関与しているが、後者は前者に比べて線虫の保持数は著しく低く、関与は部分的と言われる。属名の通り両種とも長い触角(ひげ)を持つが、体の方は何れも1-3cm程度と小型。色もマツノマダラは茶色の斑(まだら)模様、カラフトヒゲナガは黒色と地味である。アメリカでの媒介は同属の''M. carolinensis''など数種、スペインでは''M. galloprovincialis'' という種が主に関わっているという。 |
|||
==== その他の類縁線虫 ==== |
|||
これらのカミキリの成虫は特に弱った木や枯死したばかりの木に好んで産卵し、幼虫はマツ(正確にはマツ属以外のマツ科植物も食べる)の材を食べて育つ。産卵は晩春から夏にかけて行われる。幼虫は初め樹皮直下の組織を食べるが、冬になるとより内部に潜り込む。冬になるまでに一通りの成長を終えた幼虫はさなぎを経て、6月-7月にかけて羽化する。カラフトヒゲナガの場合はもう少し早いという。成虫の餌はマツの若い枝であり、材から脱出するとこれを食べに向かう(昆虫学の用語では羽化後の摂食を後食と言う)。後食をしていくうちに性成熟したカミキリは繁殖相手を求め始め、やがて交尾・産卵し夏の終わりには姿を消す。 |
|||
線虫という生き物は非常に多種類が知られ、土壌中で腐植を食べるような物から昆虫や動物に寄生して病気を引き起こすものまでいろいろである。''Burusaphelenchus''属線虫は生態面で植物寄生性であり、媒介を昆虫に頼るのを特徴としている。現在100種以上が知られており、大きく3グループに分けられるという<ref name="神崎・竹本(2012)">神崎菜摘, 竹本周平(2012)、「[https://doi.org/10.4005/jjfs.94.299 ''Bursaphelenchus''属線虫の植物病原性と媒介者の生活史特性の関連]」 『日本森林学会誌』 2012年 94巻 6号 p.299-306, {{doi|10.4005/jjfs.94.299}}, 日本森林学会</ref>。日本においてはマツノザイセンチュウ、ニセマツノザイセンチュウの他にも、[[ウコギ科]]樹木に付くタラノザイセンチュウ (''Burusaphelenchus luxuriosae'')、[[クワ]]に付くクワノザイセンチュウ (''B.conicaudatus'')などが知られる。宿主に対する病原性は一般的に低く、何らかの原因で衰弱していたものを枯死に追い込むことが出来る程度のものが多いとされる。マツ属枯れ木で見つかる''B. doui''など病原性は無いのではないかとも言われる種もある。しかしながら、中には高い病原性を持つものも存在し、特に有名なのが本病の''B. xylophilus''と[[中南米]]や[[カリブ海]]諸国の[[ヤシ]]類[[プランテーション]]で赤輪病(英名: red ring disease)を引き起こし大きな被害を出している''B. cocophilus''の2種。ただし、これら2種共に被害を出しているのは土着の植物ではなく持ち込まれた植物に対してである。最近では他の種でも病原性が比較的強いものも報告されており、マツ類苗木に病原性を示した''B. sexdentati''などがある<ref>G. Skarmoutsos andH. Michalopoulos-Skarmoutsos (2000)[https://doi.org/10.1046/j.1439-0329.2000.00198.x Pathogenicity of ''Bursaphelenchus sexdentati'', ''Bursaphelenchus leoni'' and ''Bursaphelenchus hellenicus'' on European pine seedlings.] Forest pathology30(3) 149-156, {{doi|10.1046/j.1439-0329.2000.00198.x}}</ref>。 |
|||
=== 媒介昆虫 === |
|||
マツノザイセンチュウが発見されたのに続き、この線虫がマツノマダラカミキリ(''Monochamus alternatus'')というカミキリムシによって媒介されて感染拡大を引き起こしていることが発見報告された<ref name="森本桂・岩崎厚(1972)"/>。このカミキリムシはマツ枯れ被害木からよく見つかることから以前から「松くい虫」候補の一つとして名前が挙がっていた昆虫である。 |
|||
マツノマダラカミキリはヒゲナガカミキリ属(''[[:en:Monochamus|Monochamus]]'')に属する小型のカミキリムシで、日本ではこの種以外にも近縁種が複数分布する。マツ枯れ被害にはマツノマダラカミキリ(''M. alternatus'')の他に同属のカラフトヒゲナガカミキリ(''M. saltuaris'')も関与していることが報告されているが、後者は前者に比べて線虫の保持数は著しく低く、関与は部分的と言われる。属名の通り両種とも長い触角(ひげ)を持つが、体の方は何れも1-3cm程度と小型。色もマツノマダラは茶色の斑(まだら)模様、カラフトヒゲナガは黒色と地味である。アメリカでの媒介は同属の''M. carolinensis''など数種、スペインでは''M. galloprovincialis'' という種が主に関わっているという。マツノザイセンチュウの媒介では脇役のカラフトヒゲナガカミキリではあるが、ニセマツノザイセンチュウの媒介では主役であり主要な媒介昆虫であるとされる<ref>軸丸祥大(1996)、「[https://doi.org/10.15027/690 カラフトヒゲナガカミキリの個体群動態とニセマツノザインチュウの伝播に関する研究 : 冷涼な地域におけるマツ材線虫病激害化の阻害過程]」 『広島大学総合科学部紀要. IV, 理系編』 1996年 22巻 p.191-193, {{NCID|AN10435936}}, 広島大学総合科学部</ref>。 |
|||
これらのカミキリの成虫は特に弱った木や枯死したばかりの木に好んで産卵し、幼虫はマツ(正確にはマツ属以外のマツ科植物も食べる)の材を食べて育つ。産卵は晩春から夏にかけて行われる。幼虫は初め樹皮直下の組織を食べるが、冬になるとより内部に潜り込む。冬になるまでに一通りの成長を終えた幼虫はさなぎを経て、6月-7月にかけて羽化する。カラフトヒゲナガの場合はもう少し早いという<ref>越智鬼志夫(1969)、「[https://doi.org/10.11519/jjfs1953.51.7_188 マツ類を加害するカミキリムシ類の生態 (II) ''Monochamus''属2種成虫の羽化と産卵習性などについて]」 『日本林学会誌』 1969年 51巻 7号 p.188-192, {{doi|10.11519/jjfs1953.51.7_188}}, 日本森林学会</ref>。成虫の餌はマツの若い枝であり、材から脱出するとこれを食べに向かう(昆虫学の用語では羽化後の摂食を後食と言う)。後食をしていくうちに性成熟したカミキリは繁殖相手を求め始め、やがて交尾・産卵し夏の終わりには姿を消す。 |
|||
マツ枯れ被害が確認されていない北海道においてもシラフヨツボシヒゲナガカミキリ(''M. urussovii'')という近縁種が分布する。マツ属樹木に乏しい北海道の環境のためか、この種が餌として利用するのは[[マツ科]][[トウヒ属]](''Picea'')の[[エゾマツ]](''Picea jezoensis'')や[[アカエゾマツ]](''Picea glehnii'')、同科[[モミ属]](''Abies'')の[[トドマツ]](''Abies sachalinensis'')だという。このカミキリはこれら樹木の害虫として認識されているが、線虫が北海道に侵入した場合に線虫を媒介するのかどうかについてはよくわかっていない。ただし、この種はニセマツノザイセンチュウをエゾマツやトドマツに媒介することは確認されている<ref>Togashi K., Taga Y., Iguchi, K., Aikawa T. (2008) [https://doi.org/10.1007/s10310-007-0057-1 ''Bursaphelenchus mucronatus'' (Nematoda : Aphelenchoididae) vectored by ''Monochamus urussovi ''(Coleoptera : Cerambycidae) in Hokkaido, Japan.] Journal of Forest Research 13(2), 127-131, {{doi|10.1007/s10310-007-0057-1}}</ref>。 |
|||
他の''Bursaphelenchus''属線虫を媒介する昆虫には[[カミキリムシ]]、[[キクイムシ]]、[[ゾウムシ]]、[[クワガタムシ]]、ある種のハナバチ等が知られるという<ref name="神崎・竹本(2012)"/>。なお、マツノザイセンチュウに関しては近年根の罹病木と健全木の癒合部分からも感染することが明らかになっている<ref>田中一二三, 玉泉幸一郎, 保坂武宣, (2008)、「[https://doi.org/10.11519/jfsc.119.0.119.0 根系癒合によるマツ材線虫病被害木の拡大]」 『日本森林学会大会発表データベース』 第119回日本森林学会大会 セッションID: A09, {{doi|10.11519/jfsc.119.0.119.0}}, 日本森林学会</ref>。 |
|||
<gallery> |
<gallery> |
||
| 47行目: | 61行目: | ||
File:Monochamus galloprovincialis male up.JPG|ヨーロッパ産近縁種''M. galloprovincialis'' |
File:Monochamus galloprovincialis male up.JPG|ヨーロッパ産近縁種''M. galloprovincialis'' |
||
</gallery> |
</gallery> |
||
=== 青変菌 === |
=== 青変菌 === |
||
青変菌は名前の通り木材を青く変色させることで知られる。マツ枯れによる枯死木を伐採すると切り口が青く変色していることが多く、かつては青変菌がマツ枯れの主原因ではないかとも言われていたこともある。 |
青変菌(英名:blue stain fungi)は名前の通り木材を青く変色させることで知られる菌類で''Ceratocystis''属、''Grosmannia''属、''Ophiostoma''属など複数の属に跨って知られる。マツ枯れによる枯死木を伐採すると切り口が青く変色していることが多く、かつては青変菌がマツ枯れの主原因ではないかとも言われていたこともある。日本では青変菌は木材の経済的価値を落とす菌として有名だが、世界的にみると欧米の[[ニレ属]]樹木に壊滅的被害を出し続けている[[ニレ立枯病]](オランダニレ病)を引き起こす''Ophiostoma ulmi''やアメリカのナラ類のoak wilt病(和名未定)を引き起こす''Ceratocystis fagacearum''、アメリカ西部のキクイムシの一種(''Dendroctonus ponderosae'')が引き起こすコントルタマツ(''Pinus contorta'')の病気などのように病原性の強いものも複数知られる。しかしマツ枯れの場合マツが枯死してから青変菌が増殖することなどから主たる原因ではないとされた<ref>黒田慶子, 伊藤進一郎(1992)、「[https://doi.org/10.11519/jjfs1953.74.5_383 クロマツに侵入後のマツノザイセンチュウの動きとその他の微生物相の変遷]」 『日本林学会誌』 1992年 74巻 5号 p.383-389, {{doi|10.11519/jjfs1953.74.5_383}}, 日本森林学会</ref>。マツ枯れ材に青変菌が見られるのは主にカミキリムシやキクイムシが体に付けて持ち込むからだという。 |
||
<gallery> |
|||
File:Blue stain fungus 01.jpg|青変菌の繁殖した樹木の切断面、マツ枯れ枯死木もこのようになることが多い |
|||
File:Dutch Elm Disease affecting a mature English Elm at Wst Point, NY June 2010.jpg|世界的に有名なニレ立枯病(オランダニレ病) |
|||
File:Oak wilt aerial photo.jpg|アメリカのoak wilt病(和名未定) |
|||
File:Beetle kill forest colorado.jpg|キクイムシと青変菌により枯死した大量のコントルタマツ. アメリカ・[[コロラド州]] |
|||
</gallery> |
|||
=== その他の原因説 === |
|||
線虫は[[コッホの原則]]を満たし大規模なマツ枯れを引き起こしている原因であることはほぼ間違いないが、原因・主因は他にあるとする見方も少ないながら存在する。線虫以外の病原体説、大気汚染説、酸性雨説、土壌の富栄養化によるマツの根の衰弱説などが特に有名。これらの要因でマツが衰弱し、時として枯死に至ることは確かにあり<ref>作山健 (1985) マツつちくらげ病の発生生態と防除. 岩手県林業試験場報告18号</ref><ref>佐藤邦彦, 横沢良憲, 庄司次男(1974)、「[https://agriknowledge.affrc.go.jp/RN/2010101542 マツ類の群状枯死を起こす「つちくらげ」病に関する研究(「まつくいむしによるマツ類の枯損防止に関する研究」の一部)]」『林業試験場研究報告』1974年 268号 p.13-48, 森林総合研究所</ref><ref>中村克典, 秋葉満輝, 相川拓也, 小坂肇, 伊禮英毅, 喜友名朝次 (2010)、「[https://doi.org/10.4005/jjfs.92.45 沖縄県宮古島のリュウキュウマツ枯死木およびマツノマダラカミキリからの''Bursaphelenchus''属線虫検出調査]」 『日本森林学会誌』 2010年 92巻 1号 p.45-49, {{doi|10.4005/jjfs.92.45}}, 日本森林学会</ref>、実際の山林のマツ枯れ被害には線虫を主因にこれらの要因を加えた複合的な問題が存在すると考えられている。 |
|||
<gallery> |
|||
File:Rhizina undulata.JPG|キノコの一種[[ツチクラゲ]]はマツを枯らすことがある |
|||
File:Armillaria mellea, Honey Fungus, UK 1.jpg|[[ナラタケ]]類もマツを枯らすことがある |
|||
File:Acid rain woods1.JPG|酸性雨で枯死した樹木。[[チェコ]] |
|||
</gallery> |
|||
== 発病に関する因子 == |
== 発病に関する因子 == |
||
=== マツの種類と感受性 === |
=== マツの種類と感受性 === |
||
マツの種類により線虫への感受性に差がある。日本産種は何れも |
マツの種類により線虫への感受性に差がある。日本産種は何れも感受性が高く、特に[[クロマツ]](''Pinus thunbergii'')や[[チョウセンゴヨウ]](''P. koraiensis'')は弱い。線虫が天然分布する北米産種は特に東部に分布する種を中心に線虫に抵抗性を示すものが多い。興味深いことにアジア産種であっても抵抗性を示す種がいくつか知られる、逆に北米種であっても西部やメキシコに分布するものを中心に弱いものもある。以下、古野(1982)<ref>古野東洲(1982)、「[https://hdl.handle.net/2433/191761 外国産マツ属の虫害に関する研究 : 第7報 マツノザイセンチュウにより枯死したマツ属について]」 『京都大学農学部演習林報告』 1982年 54巻 p.16-30, {{hdl|2433/191761}}, 京都大学農学部附属演習林</ref>における野外での観察による結果を基に、二井・古野(1979)<ref>二井一禎, 古野東洲(1979)、「[https://hdl.handle.net/2433/191693 マツノザイセンチュウに対するマツ属の抵抗性]」 『京都大学農学部演習林報告』 1979年 51巻 p.23-36, {{hdl|/2433/191693}}, 京都大学農学部附属演習林</ref>、や清原・徳重(1971)<ref name="マツ生立木の対する線虫接種試験"/>も加えて感受性についてのリストを示す。人工的な線虫接種試験とカミキリの嗜好が加わる野外観察では結果が変わることが予想される。また、線虫接種試験では接種時の樹齢や接種量などにより差があり、抵抗性種であっても苗木に接種したり、過剰に接種すると枯れる場合がある。ゆえに抵抗性の強弱は研究者によって多少の相違があるので参考程度にされたい。 |
||
種名の後はその種の主な分布地を示す、米東=アメリカ大陸東部、米西=アメリカ大陸西部・メキシコ、亜=アジア、欧=ヨーロッパ |
種名の後はその種の主な分布地を示す、米東=アメリカ大陸東部、米西=アメリカ大陸西部・メキシコ、亜=アジア、欧=ヨーロッパ |
||
;強抵抗性 |
;強抵抗性 |
||
:[[テーダマツ]](''Pinus taeda''、米東)、[[リギダマツ]](''P. rigida''、米東)、[[ダイオウマツ]](''P. palustris''、米東)、[[スラッシュマツ]](''P. elliotti''、米東)、[[ニイタカアカマツ]](''P. taiwanensis''、亜) |
:[[テーダマツ]](''Pinus taeda''、米東)、[[リギダマツ]](''P. rigida''、米東)、[[ダイオウマツ]](''P. palustris''、米東)、[[スラッシュマツ]](''P. elliotti''、米東)、[[ニイタカアカマツ]](''P. taiwanensis''、亜) |
||
;やや抵抗性 |
;やや抵抗性 |
||
:[[ストローブマツ]](''P. strobus''、米東) |
:[[ストローブマツ]](''P. strobus''、米東)、[[バンクスマツ]](''P. banksiana''、米東)、[[コントルタマツ]](''P. contorta''、米西)、[[バビショウ]](タイワンアカマツ, ''P. massoniana''、亜)、[[レジノーサマツ]](''P. resinosa''、米東)、クロマツ×バビショウ[[雑種第一代]](F1)、''P. tabuliformis''(亜) |
||
;やや感受性 |
;やや感受性 |
||
:[[シロマツ]](''P. bungeana''、亜)、[[モンチコラマツ]](''P. monticola''、米西)、''P. storobiformis''(米西)、[[ゴヨウマツ]](''P. parviflora''、亜)、[[アカマツ]](''P. densiflora''、亜)、[[フランスカイガンショウ]](''P. pinaster''、欧)、[[ヨーロッパアカマツ]](''P. sylvestris''、欧亜)、[[ポンデローサマツ]](''P. ponderosa''、米西)、[[ラジアータマツ]](''P. radiata''、米西)、 |
:[[シロマツ]](''P. bungeana''、亜)、[[モンチコラマツ]](''P. monticola''、米西)、''P. storobiformis''(米西)、[[ゴヨウマツ]](''P. parviflora''、亜)、[[アカマツ]](''P. densiflora''、亜)、[[フランスカイガンショウ]](''P. pinaster''、欧)、[[ヨーロッパアカマツ]](''P. sylvestris''、欧亜)、[[ポンデローサマツ]](''P. ponderosa''、米西)、[[ラジアータマツ]](''P. radiata''、米西)、 |
||
;強感受性 |
;強感受性 |
||
:[[クロマツ]](''P. thunbergii''、亜)、[[チョウセンゴヨウ]](''P. koraiensis''、亜)、[[リュウキュウマツ]](''P. luchuensis''、亜)、[[ヨーロッパクロマツ]](''P. nigra''、欧)、[[モンタナマツ]](''P. mugo''、欧)、''P. muricata''(米西)、''P. leiophylla''(米西) |
:[[クロマツ]](''P. thunbergii''、亜)、[[チョウセンゴヨウ]](''P. koraiensis''、亜)、[[リュウキュウマツ]](''P. luchuensis''、亜)、[[ヨーロッパクロマツ]](''P. nigra''、欧)、[[モンタナマツ]](''P. mugo''、欧)、''P. muricata''(米西)、''P. leiophylla''(米西) |
||
このリストには載せてないが[[ハイマツ]](''P. pumila'')や[[屋久島]]・[[種子島]]に分布する絶滅危惧種[[ヤクタネゴヨウ]](アマミゴヨウ、''P. amamiana'')<ref> |
このリストには載せてないが[[ハイマツ]](''P. pumila'')や[[屋久島]]・[[種子島]]に分布する絶滅危惧種[[ヤクタネゴヨウ]](アマミゴヨウ、''P. amamiana'')<ref>Akiba M., Nakamura K., (2005) Susceptibility of adult trees of the endangered species ''Pinus amamiana'' var. ''amamiana'' to pine wilt disease in the field. Journal of forest research 10(1): 3-7.</ref><ref>金谷整一l, 中村克典, 秋葉満輝 ほか(2005)、「[https://doi.org/10.18960/hozen.10.1_77 種子島木成国有林におけるマツ材線虫病で枯死したヤクタネゴヨウの伐倒駆除]」 『保全生態学研究』 2005年 10巻 1号 p.77-84, {{naid|110001888705}}, {{doi|10.18960/hozen.10.1_77}}, 日本生態学会</ref>も感受性である。ただし、ハイマツなどは比較的寒冷地に分布するために実験室以外の野外で影響を受けることは今のところあまりないとされる。 |
||
また、マツ属以外の[[マツ科]]樹木のうち[[モミ属]](''Abies'')、[[トウヒ属]](''Picea'')<ref>[http://www2.pref.iwate.jp/~hp1017/kenkyu/matsukuimushi.htm 岩手県林業技術センター(2005)ドイツトウヒに発生したマツ材線虫病. 林業技術情報No.21]</ref>、[[カラマツ属]](''Larix'')、[[ヒマラヤスギ属]](''Cydrus'')の一部樹木についてマツノザイセンチュウによる枯死例が報告されている。 |
また、マツ属以外の[[マツ科]]樹木のうち[[モミ属]](''Abies'')、[[トウヒ属]](''Picea'')<ref>[http://www2.pref.iwate.jp/~hp1017/kenkyu/matsukuimushi.htm 岩手県林業技術センター(2005)ドイツトウヒに発生したマツ材線虫病. 林業技術情報No.21]</ref>、[[カラマツ属]](''Larix'')、[[ヒマラヤスギ属]](''Cydrus'')の一部樹木についてマツノザイセンチュウによる枯死例が報告されている。 |
||
| 80行目: | 111行目: | ||
マツ林の放棄に伴う土壌の富栄養化も間接的誘因となることが以前より示唆されている。健全なマツは吸収根のほとんどが[[外菌根]]となっており、それにより養分・水分の乏しい環境に適応しているとされているが、土壌の富栄養化によって吸収根は土壌表層に集まり、なおかつ菌根の発達が悪くなる。土壌の表層は夏の乾燥の影響を受けやすい部分でもあり、このような状態では水ストレスに敏感になると考えられている。 |
マツ林の放棄に伴う土壌の富栄養化も間接的誘因となることが以前より示唆されている。健全なマツは吸収根のほとんどが[[外菌根]]となっており、それにより養分・水分の乏しい環境に適応しているとされているが、土壌の富栄養化によって吸収根は土壌表層に集まり、なおかつ菌根の発達が悪くなる。土壌の表層は夏の乾燥の影響を受けやすい部分でもあり、このような状態では水ストレスに敏感になると考えられている。 |
||
== 松枯れの蔓延とその影響 == |
|||
=== 生態面 === |
|||
マツ枯れの蔓延によってクロマツ林が壊滅し、広葉樹が侵入してきた海岸の事例では上層の構成種の変化だけでなく、下層の植生や土壌成分に変化があったことが報告されている。<ref>藤田恵美, 中田誠 (2001)、「[https://doi.org/10.11519/jjfs1953.83.2_84 海岸砂丘地のクロマツ林における広葉樹の混交による立地環境の変化 新潟県下越地方の事例]」 『日本森林学会誌』 2001年 83巻 2号 p.84-92, {{doi|10.11519/jjfs1953.83.2_84}}</ref>。 |
|||
=== 人間生活 === |
|||
== 日本における防除方法とそれに対する批判 == |
== 日本における防除方法とそれに対する批判 == |
||
| 86行目: | 123行目: | ||
==== カミキリムシの駆除 ==== |
==== カミキリムシの駆除 ==== |
||
[[File:Beauveria.jpg|''Beauveria bassiana''は多犯性で様々な宿主に感染する。これはバッタだが、カミキリも同じ菌が殺す|300px|thumb]] |
|||
線虫を媒介するカミキリムシを駆除し材線虫病の拡大を防ぐ方法、幼虫・成虫それぞれに方法がある。 |
線虫を媒介するカミキリムシを駆除し材線虫病の拡大を防ぐ方法、幼虫・成虫それぞれに方法がある。 |
||
幼虫を標的としたものとしては、カミキリが衰弱状態や枯死したばかりのマツに好んで産卵する性質を利用し、秋から翌年春にかけて枯死したばかりの木を伐採する方法がある。その後伐採木の焼却、粉砕、もしくは薬剤 |
幼虫を標的としたものとしては、カミキリが衰弱状態や枯死したばかりのマツに好んで産卵する性質を利用し、秋から翌年春にかけて枯死したばかりの木を伐採する方法がある。その後伐採木の焼却、粉砕、もしくは薬剤散布によって材の中のカミキリ幼虫を殺す。なお、薬剤散布の場合は幼虫が材内部に食い進む冬以前に行わないと殺虫効率が悪いという。これらは「伐倒駆除」と呼ばれ重労働である。近年薬剤散布方法が見直され、被害木を適当な大きさに玉切りしたあと薬剤散布とビニールシートでの被覆をセットで行うようになってきており、単なる散布と言うより燻蒸に近くなった。この方法の利点として薬剤の飛散がなく殺虫効率も高いこと、さらに破砕や焼却といった作業に比べてその場で処理できることが挙げられる。薬剤には有機リン化合物や[[カルバミン酸]]系化合物(通称カーバム剤・カーバメート剤、商品名NCS等)などが使われる。 |
||
成虫を標的にしたものとしては成虫の羽化してくる初夏(5-7月)に、カミキリが後食するマツの若い枝がある樹冠部に殺虫剤を散布し、カミキリを殺すことで線虫の感染を防ぐというものである。後食の開始と線虫がカミキリから脱出しマツに感染するのには僅かではあるがタイムラグがあるのでこの間にカミキリを殺すことは感染防止に有効とされる。主に大型噴霧器による地上からの散布か[[ヘリコプター]]による空中散布が採られる |
成虫を標的にしたものとしては成虫の羽化してくる初夏(5-7月)に、カミキリが後食するマツの若い枝がある樹冠部に殺虫剤を散布し、カミキリを殺すことで線虫の感染を防ぐというものである。後食の開始と線虫がカミキリから脱出しマツに感染するのには僅かではあるがタイムラグがあるので<ref name="森本桂・岩崎厚(1972)"/>この間にカミキリを殺すことは感染防止に有効とされる。主に大型噴霧器による地上からの散布か[[ヘリコプター]]による空中散布が採られる。薬剤には後述のように農薬の定番[[有機リン化合物|有機リン系]]か、最近はそれに比べて人畜毒性の低いと言われる[[ネオニコチノイド]]系が使われる。散布には周辺の[[ドリフト (農薬)|ドリフト被害]]などからの批判もある。また、農薬の種類によっては[[森林認証制度]]を利用できなくなる場合がある。最近では薬の他に[[昆虫病原糸状菌]]、特にその中でも''Beauveria''属菌を使う殺虫方法なども提案されている。ただし、この病原菌も農薬同様非選択性的性格が強く様々な昆虫を殺してしまう。 |
||
カミキリ特に幼虫時期を対象とした天敵動物による防除、いわゆる[[生物農薬]]についても研究が行われているが、広く実用化されたものは無い。鳥類ではキツツキの一種の[[アカゲラ]]<ref>中村充博, 鈴木祥悟(2007)、「[http://id.nii.ac.jp/1318/00001336/ アカゲラ誘致のための人工巣丸太の架設実験]」 『総合政策』 2007年 9巻 1号 p.49-58, 森林総合研究所東北支所</ref>、寄生性・捕食性昆虫として[[ホソカタムシ]]類<ref>岡本安順(1999)、「[https://doi.org/10.20660/applfor.8.0_229 マツノマダラカミキリの天敵昆虫サビマダラオオホソカタムシの寄生状況と生態調査]」 『森林応用研究』 1999年 8巻 p.229-232, {{doi|10.20660/applfor.8.0_229}}, 応用森林学会</ref><ref>喜友名朝次(2009)、「[https://agriknowledge.affrc.go.jp/RN/2010813283 松くい虫天敵昆虫防除技術開発研究]」『沖縄県森林資源研究センター研究報告』 2011年 52号 p.12-15, {{naid|120005321945}}, 沖縄県森林資源研究センター</ref><ref>牧本卓史(2011)、「[https://agriknowledge.affrc.go.jp/RN/2010831525 サビマダラオオホソカタムシの松くい虫防除への適用]」 『岡山県農林水産総合センター研究報告』 2011年 27号 p.73-82, {{naid|120005330890}}, 岡山県農林水産総合センター森林研究所</ref>、[[コクヌスト]]類<ref>上田明良, 藤田和幸, 浦野忠久, 山田倫章(1999)、「[https://doi.org/10.20660/applfor.8.0_169 オオコクヌスト成虫放虫によるマツノマダラカミキリ捕食実験]」 『森林応用研究』 1999年 8巻 p.169-172, {{doi|10.20660/applfor.8.0_169}}, 応用森林学会</ref>、[[寄生蜂]]<ref>岡本安順(1997)、「[https://doi.org/10.20660/applfor.6.0_131 スギカミキリ・マツノマダラカミキリに対するクロアリガタバチの寄生実験]」 『森林応用研究』 1997年 6巻 p.131-134, {{doi|10.20660/applfor.6.0_131}}, 応用森林学会</ref>等が注目されており、生息密度を上げることや効率的な捕食・寄生を行う方法などが研究されている。線虫の移動を助けるカミキリではあるが、ある種の線虫はカミキリに寄生する。マツノマダラカミキリから見つかった線虫は''Contortylenchus genitalicola''として新種記載され<ref>H. Kosaka, and N. Ogura, (1993) "[https://doi.org/10.1303/aez.28.423 ''Contortylenchus genitalicola'' n.sp.(Tylenchida:Allantonematidae)from the Japanese Pine Sawyer, ''Monochamus alternatus''(Coleoptera:Cerambycidae)]". 『日本応用動物昆虫学会誌』 1993年 28巻 4号 P.423-432, 日本応用動物昆虫学会</ref>、しかも卵巣に寄生するということから不妊化効果が期待されたが、完全な不妊雌成虫を作り出すのは難しくこの計画はとん挫した。なお、海外の事例ではオセアニアにおける北米原産マツ類に枯死被害をもたらす侵入昆虫[[ノクチリオキバチ]](''Sirex noctilio'')においてキバチ雌成虫を不妊化する寄生線虫が見つかっており<ref>Bedding R. A. (1968) ''Deladenus wilsoni'' n. sp. and ''D. siricidicola'' n. sp. (Neotylenchidae), entomophagous-mycetophagous nematodes parasitic in siricid woodwasps. Nematologica 14: 515-525.</ref>、この線虫を用いた被害のコントロールも実現している<ref>Bedding, R.A., and Akhurst R. J., (1971)Use of the nematode ''Deladenus siridicola'' in the biological control of ''Sirex noctilio'' in Australia. Australian Journal of Entomology 13: 129-135.</ref>。また、キバチの線虫とは属が異なるが、日本においても[[スズメバチ]]女王蜂を不妊化する寄生線虫が見つかっている<ref>Sayama K., Kosaka H., and Makino, S., (2007) The first record of infection and sterilization by the nematode ''Sphaerularia'' in hornets (Hymenoptera, Vespdiae, Vespa). Insectes Sociaux54: 53-55.</ref>。 |
|||
特定の林分でマツ枯れを防除する場合、侵入初期に異常を示した少数のマツを的確に伐倒処理すれば、林内で感染サイクルが回って被害が拡大するのを防ぐことができる。しかし近隣に激害状態の林分があれば、そこから線虫保持カミキリが侵入することは防げない。林内でカミキリが発生して激害とならないよう薬剤散布などの手法も併用して駆除しつつカミキリの飛来がなくなるまで持ちこたえることができれば、防除は成功する。[[鹿児島県]]の[[吹上浜]]や[[佐賀県]]の[[虹ノ松原]]はそのようにして防除に成功した例である。 |
|||
寄生線虫に代わって注目されているものに、[[ボルバキア]]がある。ボルバキアは昆虫や線虫に寄生する細菌で宿主雌成虫の産卵により次世代へと繋がるために、宿主昆虫の生殖機構を乗っ取り宿主雌個体、ひいては自分に有利に利用することが多い。他にも犬のフィラリア症では[[フィラリア]]に感染したボルバキアが病原性を高めたり、ある系統のボルバキアが感染した[[ネッタイシマカ]]は[[デング熱]]を媒介しにくくなる、宿主昆虫の寿命を縮めるなどの面白い性質が知られるものもある。近年マツノマダラカミキリを調べた結果、ボルバキアは単なる感染ではなくカミキリの常染色体に組み込まれているといい、[[遺伝子の水平伝播]]を起こしたことが示された<ref>相川拓也(2012)、「[https://doi.org/10.4005/jjfs.94.292 マツノマダラカミキリに残る共生細菌感染の痕跡]」『日本森林学会誌』 2012年 94巻 6号 p.292-298, {{DOI|10.4005/jjfs.94.292}}, 日本森林学会</ref>。昆虫のような高等生物でこのような水平伝播は珍しいという。ボルバキアは前述のように宿主の生殖機構のコントロール能力を持つことがあるためカミキリについても[[不妊虫放飼]]のような効果があるのではないかという仮説の基で研究が進められている。 |
|||
発病後一気に枯死に至らず、年が明けた春先、場合によっては夏近くまで生存する個体が稀にある。カミキリ成虫が羽化する時期にこれらの衰弱した個体がある場合、マツが[[アレロケミカル|カイロモン]]となるエタノールや[[テルペン]]類を発して性成熟したカミキリを誘引するため、防除の障害となる。 |
|||
他にはマツが弱った時に発する匂いを模したもので、産卵期のカミキリを誘因する薬剤などもある。マツノマダラカミキリでの誘因調査事例では比較的上の方によく集まるという<ref>上田明良(1998)、「[https://doi.org/10.20660/applfor.7.0_109 マダラコールを用いた誘因トラップの設置高別捕獲調査]」 『森林応用研究』 1998年 7巻 p.109-112, {{doi|10.20660/applfor.7.0_109}}, 応用森林学会</ref>。マツノマダラカミキリ成虫が忌避行動をとる[[周波数]]の音波があることが発見され、農薬に変わる新たな防除手段になるのではないかと期待されている<ref>高梨琢磨.・小島渉(2012)森林昆虫における振動情報の機能解明と害虫防除への応用. 木材保存38(4):157-162.</ref> |
|||
特定の林分でマツ枯れを防除する場合、侵入初期に異常を示した少数のマツを的確に伐倒処理すれば、林内で感染サイクルが回って被害が拡大するのを防ぐことができる。しかし近隣に激害状態の林分があれば、そこから線虫保持カミキリが侵入することは防げない。理論上は林内でカミキリが発生して激害とならないよう薬剤散布による殺虫、近隣林分の樹種転換によってカミキリの飛来が充分少なくなるまで持ちこたえることができれば、防除は成功すると考えられている。日本における完全な根絶例は[[沖永良部島]]の一例で1995年に根絶宣言がなされている。この事例では空中散布によるカミキリ駆除の効果が高かったことが指摘されている<ref>Muramoto M. ,(1999) Ending of pine wilt disease in Okinoerabu island, Kagoshima prefecture. In Futai, K., Togashi, K. & Ikeda, T. eds. Sustainability of Pine Forests in Relation to Pine Wilt and Decline. Shokado shoten, Kyoto.</ref>。この他に[[鹿児島県]]の[[吹上浜]]<ref>Nakamura K., Yoshida N., (2004) Successful control of pine wilt disease in Fukiage-hama seacoast pine forest in southwestern Japan. In Mota M., Viera P., eds. The pine wood nematode ''Bursaphelenchus xylophilus''. proceeding international workshop, University of Evora, Portugal August 2001.</ref>や[[佐賀県]]の[[虹ノ松原]]で被害をほぼ抑えることに成功している。ただし、吹上浜については2008年頃から薬剤散布が行われなくなって再び激害化してしまい、図らずも空中散布の有効性が示される結果となった。 |
|||
発病後一気に枯死に至らず、年が明けた春先、場合によっては夏近くまで生存する個体があり寒冷地では半数以上がこのタイプと言われる。カミキリ成虫が羽化する時期にこれらの衰弱した個体がある場合、マツが[[アレロケミカル|カイロモン]]となるエタノールや[[テルペン]]類を発して性成熟したカミキリを誘引するため、防除の障害となる。 |
|||
<gallery> |
|||
File:Beauveria.jpg|''Beauveria bassiana''は多犯性で様々な宿主に感染する。これはバッタだが、カミキリも同じ菌が殺す |
|||
File:Dendrocopos major 3 (Marek Szczepanek).jpg|アカゲラ |
|||
</gallery> |
|||
==== 線虫の駆除 ==== |
==== 線虫の駆除 ==== |
||
薬剤を樹幹に注入するか土壌に散布し根から吸収させる。効果では線虫増殖抑制作用のものと殺線虫作用のものに大別される。 |
|||
殺線虫剤や線虫増殖抑制剤を樹幹に注入する。材線虫病は感染後急激に発症枯死に至り、また管類の閉塞が見られることもあって、投与は線虫の感染前に予防的に行われる。防除効果は極めて高いが幹に傷を付けることへの景観的問題、薬剤費用、1-3年に一回の継続的な投与が必要などの欠点があり、広範囲に用いるのは現実的ではない。主に各地の銘木や貴重な個体群、個人所有の庭木などに対して用いられる。 |
|||
代表的なものにモランテル(酒石酸塩が商品名グリーンガード)、メスルフェンホス(商品名ネマノーン)、[[レバミゾール]](塩酸塩が商品名センチュリー等)、[[マクロライド]]系と呼ばれ、土壌中の細菌が産生する3種、ネマデクチン(商品名メガトップ)・[[エマメクチン]](商品名ショットワンツー)・ミルベクチン(商品名マツガード)などがある。これらの薬剤の有効成分の多くは人を含む動物における線虫感染症にも用いられており、中には殺虫剤として使われるものもある。その他の薬でも効くものがあり、線虫も人や昆虫と同じく[[アセチルコリン]]を[[神経伝達物質]]として使うので神経伝達作用を阻害する作用の薬、例えばアセチルコリンの速やかな分解を阻害する有機リン剤なども線虫に有害である<ref>松浦邦昭・細野隆次(1987)[https://cir.nii.ac.jp/crid/1390282679315044480 マツノザイセンチュウのアセチルコリンエステラーゼの特性と防除剤によるその活性阻害]. 日本線虫学会誌17: 17-22.</ref><ref>小倉信夫(2004)[https://cir.nii.ac.jp/crid/1050845763618468608 培養条件下でのマツノザイセンチュウの増殖に対する数種殺虫剤の抑制効果]. 日本線虫学会誌34(2): 99-102</ref>。 |
|||
マツ材線虫病は感染後急激に発症枯死に至り、また管類の閉塞が見られることもあって、投与は線虫の感染前に予防的に行われることが多い。防除効果は極めて高いがマツ1本あたり数千円の薬剤費用がかかり、かつ1-3年に一回の継続的な投与が必要だという費用的な問題の他、ドリルによって穴を開けることや傷口の不適切な処理による傷害、また薬害が出る場合もある等の欠点がある<ref>竹下努 (1992) マツ材線虫病の注入傷害防止に関する研究. 鳥取県林業試験場研究報告第34号.</ref><ref>中川茂子 (2001) [https://cir.nii.ac.jp/crid/1390005822569026304 マツ枯れ防止樹幹注入剤施用により発生する異常]. 樹木医学研究5(1): 13-20. </ref>。薬剤の成分調整や施用方法の改良によりこれらの問題は改善されつつあるが、費用面・労働面の負担が依然として農薬散布に比べ大きく広範囲に用いるのは現実的ではない。主に各地の銘木や貴重な個体群、個人所有の庭木などに対して用いられる。 |
|||
カミキリの場合同様、天敵生物の利用もいくつか研究されているが実用に至ったものは無い。マツ丸太に対して線虫と[[ヒラタケ]]等の[[線虫捕食菌]]を共に接種した結果では、マツノザイセンチュウの増殖を抑制することがしめされている<ref>真宮靖治・平塚美幸・村田政穂(2005)[https://cir.nii.ac.jp/crid/1520009408000329984 木材腐朽菌のマツノザイセンチュウに対する捕食効果.] 日本線虫学会誌 35(1): 21-30.</ref>。また[[トリコデルマ]](''Trichoderma'')属菌や[[アクレモニウム]](''Acremonium'')属菌は線虫に対して有害であるとされる<ref name="マツノザイセンチュウの生活環に関連する糸状菌(1)"/>。マツノザイセンチュウは系統によって病原性が大きく異なり、また強毒性の系統でも培養を重ねると病原性を失う<ref name="清原 (1976)"/>。この病原線虫の発見者の一人である清原友也は弱毒性系統の線虫を事前に接種しておくと、後に強毒性系統の線虫を接種した時に枯死しにくいという[[ワクチン]]のような誘導抵抗性効果がある<ref>清原友也 (1990) [https://doi.org/10.1271/nogeikagaku1924.64.1251 マツ材線虫病における誘導抵抗性]. 日本農芸化学会誌64(7): 1251-1253.</ref>という面白い現象を報告しているが、これも実用化には至っていない。 |
|||
他の植物寄生線虫による被害、特に農作物における線虫の被害は土壌病害という扱いになるものが多く、薬剤燻蒸の他にも[[連作]]を避けたり、殺線虫効果のある[[マリーゴールド]]等の植え付け、天地返しなどの伝統的対処法が知られている。細菌の一種[[バチルス・チューリンゲンシス]](''Bacillus thuringiensis'')の産生する毒素は昆虫に有害、人に無害ということで[[BT剤]]などと呼ばれ農薬に用いられてたり、毒素を作り出す遺伝子を農作物に組み込んで[[遺伝子組み換え作物]]として利用されたりしている。近年この毒素が昆虫のみならずある種の線虫にも有害であるという報告がなされており<ref>B. S. Griffiths, S. Caul, J. Thompson, A. N. E. Birch, C. Scrimgeour, M.N. Anderson, J. Corter, A. Messean, C. Sausse, B. Lacroix, P.H. Krogh, (2005) A comparison of soil microbial community structure, protozoa and nematodes in field plots of conventional and genetically modified maize expreessing the ''Bacillus thuringiensis'' Cry1Ab toxin. Plant and soil 275: 135-146. </ref>、農作物だけでなく他の分野への応用が期待される。 |
|||
<gallery> |
|||
File:ネマノーン (483707052).jpg|薬剤の樹幹注入風景 |
|||
File:Levamisole2.svg|レバミゾール |
|||
File:Emamectins.svg|エマメクチン |
|||
File:Milbemectin structure.svg|ミルベメクチン |
|||
File:Thionazin.svg|チオナジン |
|||
</gallery> |
|||
==== 被害材の移動制限 ==== |
==== 被害材の移動制限 ==== |
||
在来、外来問わずマツ枯れ被害材の安易な移動は、木材内に線虫保持カミキリがいる可能性を考えると慎むべきである。離島での発生はもちろん、そもそも日本での発生も材内にいた線虫保持カミキリから始まったと考えられている。 |
在来、外来問わずマツ枯れ被害材の安易な移動は、木材内に線虫保持カミキリがいる可能性を考えると慎むべきである。離島での発生はもちろん、そもそも日本での発生も材内にいた線虫保持カミキリから始まったと考えられている。 |
||
日本では森林病害虫等防除法第3条 |
日本では森林病害虫等防除法第3条第5項に基づき移動制限・移動禁止措置が義務付けられている<ref>{{Cite web|和書|url=https://laws.e-gov.go.jp/law/325AC0000000053#16 |title=森林病害虫等防除法(昭和二十五年法律第五十三号)第3条第5項:駆除命令、移動を制限 |website=e-Gov法令検索 |publisher=総務省行政管理局 |date=2016-05-20 |quote=2017年4月1日施行分|accessdate=2020-01-15}}</ref>が、平成の時代に入っても[[奄美大島]]など離島部で新規にマツ枯れの報告があるところを見ると守られているとは言い難いのが現状である。日本国外でも''Monochamunos''属カミキリムシの侵入を警戒している国は多く、木材輸出入の際には剥皮や乾燥を求められることが多い。 |
||
==== 予防的な在来マツの伐採、樹種の切り替え ==== |
==== 予防的な在来マツの伐採、樹種の切り替え ==== |
||
激害林分及び線虫が侵入すれば激害となることが予想される林分は、積極的に伐採して樹種転換を図り、被害拡大を食い止めることも行われている。2006年には青森県において |
激害林分及び線虫が侵入すれば激害となることが予想される林分は、積極的に伐採して樹種転換を図り、被害拡大を食い止めることも行われている。2006年には青森県においてマツ枯れの北上を阻止するために隣接する岩手・秋田の2県のうち、病気の北上の著しい[[秋田県]]との県境に約2km幅の防除帯を設けマツをすべて除去するという対策が採られた。なお、[[青森県]]内では県境から遠いにも拘らず[[蓬田村]]において2010年1月に線虫感染による枯死木が初確認され、久々に新規都道府県での感染確認となった。その後2013年には危惧されていた通り秋田県に隣接する[[深浦町]]で枯死木が確認されるなど徐々に被害が出始めている。 |
||
また、カミキリは線虫の感染に限らず弱ったマツを産卵対象として好むために、林内の競争に負けて弱った個体を間伐等で間引くのはカミキリを誘因・産卵させないために有効とされる。 |
また、カミキリは線虫の感染に限らず弱ったマツを産卵対象として好むために、林内の競争に負けて弱った個体を間伐等で間引くのはカミキリを誘因・産卵させないために有効とされる。 |
||
==== 抵抗性品種の植栽・開発 ==== |
==== 抵抗性品種の植栽・開発 ==== |
||
かつては抵抗性の北米種を植えることが行われた。テーダマツ |
かつては抵抗性の北米種を植えることが行われた。テーダマツやリギダマツが代表的である。特にテーダマツは原産地では樹高40-50mに達することもある大型種で在来アカマツと比べて苗木の成長もきわめて速い<ref>千葉春美・石井幸男・富岡甲子次・飯塚三男.(1963)赤沼におけるテーダマツの成長. 林業試験場報告161号.</ref>などと対マツ枯れ以外の面も期待されたが、地上部に比べて根の発達が貧弱で風で損傷しやすいことや、外来種問題などから現在は廃れてしまった。他にも在来のクロマツにバビショウ(タイワンアカマツ、''P. massoniana'')を掛け合わせた雑種などが開発されたことがある。バビショウはアジア産種でありながら線虫に対して強い抵抗性があり、この雑種も抵抗性を持ったという。ただし、これも線虫には抵抗性があっても他の害虫に弱いという欠点があり広い普及には至らなかった<ref>中井勇・光枝和夫・大畠誠一 (1995) [https://hdl.handle.net/2433/192091 マツ属の種間交雑からみた種の類縁関係とF1雑種の生育状況について]. 京都大学農学部演習林報告67: 1-18.</ref>。 |
||
現在主流なのは在来種の抵抗性個体の選抜である。マツ枯れ被害の大きな森林でも枯死せず生き残った個体から種子を採り、発芽させた苗木に線虫を投与して枯死しなかったものを選抜する。実生の子は親のクローンではないために必ずしも抵抗性を持つとは限らない<ref>後藤晋・宮原文彦・井出雄二(2002) クロマツ種苗のマツ材線虫病抵抗性に対する花粉親の寄与 日本林学会誌</ref>ために、線虫の接種による抵抗性の検定が欠かせず、手間と費用がかかるのが欠点である。[[挿し木]]などの無性繁殖では子供は親の形質をそのまま受け継ぐために、抵抗性系統の母樹から低コストで効率的に生産ができると考えられている。一方でマツは一般的に挿し木による繁殖が難しいことで知られており、効率的な発根・定着条件を探す研究が続けられている<ref>石川広隆(1968) マツ・カラマツ類を中心としたさし木困難樹種の不定根の形成に関する基礎研究 1968 林試研報214号</ref><ref>後藤晋 (1999) クロマツの挿し木増殖における発根条件の検討 日林九支研論52号</ref><ref> |
現在主流なのは在来種の抵抗性個体の選抜である。マツ枯れ被害の大きな森林でも枯死せず生き残った個体から種子を採り、発芽させた苗木に線虫を投与して枯死しなかったものを選抜する。実生の子は親のクローンではないために必ずしも抵抗性を持つとは限らない<ref>後藤晋・宮原文彦・井出雄二(2002) クロマツ種苗のマツ材線虫病抵抗性に対する花粉親の寄与 日本林学会誌</ref>ために、線虫の接種による抵抗性の検定が欠かせず、手間と費用がかかるのが欠点である。[[挿し木]]などの無性繁殖では子供は親の形質をそのまま受け継ぐために、抵抗性系統の母樹から低コストで効率的に生産ができると考えられている。一方でマツは一般的に挿し木による繁殖が難しいことで知られており、効率的な発根・定着条件を探す研究が続けられている<ref>石川広隆(1968) マツ・カラマツ類を中心としたさし木困難樹種の不定根の形成に関する基礎研究 1968 林試研報214号</ref><ref>後藤晋 (1999) クロマツの挿し木増殖における発根条件の検討 日林九支研論52号</ref><ref>松永孝治・大平峰子・倉本哲嗣(2009) [https://cir.nii.ac.jp/crid/1390001205228692736 さし穂サイズと採穂台木の形態的要因がクロマツさし木苗の生産効率に与える影響].日本森林学会誌91(5): 335-343.</ref>、近年の研究では発根率の良い若い苗木時代に植物ホルモンの[[サイトカイニン]]を投与してやると発根率の良い挿し穂が量産できるとされている。アカマツには抵抗性系統が比較的多く見つかっているが、クロマツの抵抗性系統は少ない。また、これらについても抵抗性はあるが、樹形や成長速度などの面は考慮されておらず課題が残る。 |
||
病理的に不明な部分があること、日本人の[[遺伝子組み換え作物]]に対する嫌悪感もあってか遺伝子組み換えの抵抗性マツを作ろうという動きは殆どない。海外種との交雑実験も前述のように廃れてしまった。 |
|||
=== 防除事業への批判 === |
=== 防除事業への批判 === |
||
現状の松くい虫防除事業には、一定の批判がある。 |
現状の松くい虫防除事業には、一定の批判がある。 |
||
カミキリ成虫を標的にした農薬の空中散布は根強い反対意見がある。農薬散布は1977年(昭和52年)4月18日付法律第18号の松くい虫被害対策特別措置法 <!--<ref>[https://web.archive.org/web/20161011180158/http://law.e-gov.go.jp/haishi/S52HO018.html 松くい虫被害対策特別措置法] 電子政府の総合窓口e-gov</ref>-->で制度化された。(その後、1997年(平成9年)の特別措置法の失効に伴い、森林病害虫等防除法が改正され必要なものについては、この法律に取り込まれている。同第七条の二、防除実施基準など)カミキリ駆除を目的として散布される農薬には大きく分けて[[有機リン]]系と[[ネオニコチノイド]]系があり、何れも昆虫の神経に作用する。有機リン系化合物は現在の農薬の定番とも言える薬剤ではあるが[[サリン]]などの[[化学兵器]]にもなる猛毒ガスも知られ、ネオニコチノイドは[[ミツバチ]]の異常行動の原因として疑われるなど何れも印象はあまり良くない。前者には[[フェニトロチオン]](商品名MEP剤、スミチオン、スミパイン)や[[マラチオン]](商品名マラソン)、プロチオホス(商品名プロチオン、ソビーT)。後者には[[アセタミプリド]](商品名マツグリーン)やチアクロプリド(商品名エコワン、エコファイター)などがある。これらの農薬はカミキリ以外の昆虫にも有害(所謂「非選択性」の農薬)であるため、生物多様性などに悪影響を与えるのではと言う声もある。 |
|||
まず、クロマツ、アカマツは、いずれも広葉樹の高木の生育が困難な厳しい環境条件下で局所的に安定した群落を維持することを除くと、広域で安定した[[森林]]を作らない。むしろ、[[遷移 (生物学)|植物遷移]]の上では、[[裸地]]に定着する先駆者樹木であり、林内が暗く富栄養になると次世代への更新ができない。そのため、松林を維持するには常に林床を貧栄養かつ明るい条件に維持する必要がある。かつて、松林の落葉落枝は広く[[燃料]]として利用され、そのために人里周辺では松林が維持されてきた。現在それが行われなくなったため、松林の減少するのは当然であり、マツ材線虫病は単にそれを加速したにすぎないという考えがある。 |
|||
有機リン・カーバメート(有機窒素)などの[[コリンエステラーゼ]]阻害剤による野生生物による影響としては以下に示すような報告がある。ただし、上に挙げたようなマツ枯れに実際に使う薬剤だけでなく広く「有機リン・カーバメート」としていることには留意されたい。たとえば野鳥については清宮ら(2007)<ref>清宮幸男・古川岳大・高橋真紀・田村貴・村上隆宏・関慶久・佐々木幸治 (2007)[https://doi.org/10.12935/jvma1951.60.191 野鳥のコリンエステラーゼ阻害剤中毒]. 日本獣医師会雑誌</ref>、水生生物は岩田(1981)<ref>岩田治郎(1981)[https://cir.nii.ac.jp/crid/1390002184874037504 農薬の沿岸生態系への影響(シンポジウム:沿岸海域における人工有機化合物汚染)]. 沿岸海洋研究ノート19(1):47-55.</ref>、菊地・若林(1997)<ref>菊地幹夫・若林明子(1997)[https://doi.org/10.2331/suisan.63.627 オオミジンコによる河川水中の化学物質の有害性モニタリング]. 日本水産学会誌. 63(4):627-633.</ref>など。 |
|||
ちなみに、同じく昆虫が病原体を媒介とする[[ニレ立枯病]]においては病原菌媒介者である[[キクイムシ]]類を標的とした[[DDT]]等の非選択性農薬の散布が行われたが、薬剤の[[生物濃縮]]による小鳥の中毒死など他生物に影響が出たので中止された。DDT禁止運動の発端になったのが[[レイチェル・カーソン]]著の有名な「[[沈黙の春]]」(原題:Silent Spring)であり、民衆を大きく動かした事例として知られる。 |
|||
<gallery> |
|||
File:Fenitrothion Structural Formulae .V.1.svg|フェニトロチオン(有機リン系殺虫剤) |
|||
File:Malathion.png|マラチオン(有機リン系) |
|||
File:Acetamiprid Structural Formulae V.1.svg|アセタミプリド(ネオニコチノイド系) |
|||
File:Thiacloprid structure.svg|チアクロプリド(ネオニコチノイド系) |
|||
File:DDT.svg|【参考】DDT(有機塩素系) |
|||
</gallery> |
|||
また、クロマツ、アカマツは、いずれも広葉樹の高木の生育が困難な厳しい環境条件下で局所的に安定した群落を維持することを除くと、広域で安定した[[森林]]を作らない。むしろ、[[遷移 (生物学)|植物遷移]]の上では、[[裸地]]に定着する先駆者樹木であり、林内が暗く富栄養になると次世代への更新ができない。そのため、松林を維持するには常に林床を貧栄養かつ明るい条件に維持する必要がある。かつて、松林の落葉落枝は広く[[燃料]]として利用され、そのために人里周辺では松林が維持されてきた。現在それが行われなくなったため、松林の減少するのは当然であり、マツ材線虫病は単にそれを加速したにすぎないという考えがある。 |
|||
この考えに立てば、必要のなくなった松林では松くい虫防除は必ずしも行う必要がなく、むしろ殺虫剤の広域散布による弊害を考慮すべきである。用いられている殺虫剤は選択的にマツノマダラカミキリを殺すものではなく、森林の[[生物群集]]を構成する[[昆虫]]など地上生の[[節足動物]]を非選択的に殺すものであるために、森林の生態系を乱し多様性を損なう。農薬の人体への健康被害の研究と比べても、地域生態系への影響評価の研究はまだ十分とは言えず、この観点からの防除対策の評価が必要である。また、枯死した松を切り倒す作業による周囲の攪乱によって、自然植生の回復が遅れたり、被害が広がったと思われる例もある。 |
この考えに立てば、必要のなくなった松林では松くい虫防除は必ずしも行う必要がなく、むしろ殺虫剤の広域散布による弊害を考慮すべきである。用いられている殺虫剤は選択的にマツノマダラカミキリを殺すものではなく、森林の[[生物群集]]を構成する[[昆虫]]など地上生の[[節足動物]]を非選択的に殺すものであるために、森林の生態系を乱し多様性を損なう。農薬の人体への健康被害の研究と比べても、地域生態系への影響評価の研究はまだ十分とは言えず、この観点からの防除対策の評価が必要である。また、枯死した松を切り倒す作業による周囲の攪乱によって、自然植生の回復が遅れたり、被害が広がったと思われる例もある。 |
||
長野県松本市では、アカマツ林の松枯れ被害のうちマツ材線虫病による被害木の主原因とされるマツ材線虫を媒介伝播する主原因とされるマツノマダラカミキリの羽化後の成虫の駆除のみを目的としたニコチノイド系殺虫剤の無人ヘリコプターによる空中散布が2013年から行われているが、その効果は限定的で2019年現在散布地のアカマツ林の松枯れはさらに拡大もしくは枯損が拡がっている。このため[[ドリフト (農薬)|ドリフト]]等による健康被害や発達障害リスク税金補助金の無駄遣いを憂う複数の弁護士及び松本市民らによる反対署名活動や市街地での反対デモ行進など大規模な空中散布反対運動が展開され、2017年から松本市を被告とした訴訟がいくつか提起されている。この問題は国会でも取り上げられ、週刊誌やネットニュース等でも報道されている。<ref>{{Cite web|和書|title=松本の松枯れを考える住民の会|url=https://www.facebook.com/MATSUMOTONONOYAKUSANPUWOKANGAERUJUMINNOKAI/posts/2079202742326092|website=www.facebook.com|accessdate=2019-05-26|language=ja}}</ref><ref>{{Cite web|和書|title=発達障害リスク「農薬」に対策遅れる日本 禁止どころか緩和も 〈週刊朝日〉|url=https://dot.asahi.com/articles/-/116898|website=AERA dot. (アエラドット)|date=2019-03-06 |accessdate=2019-05-26|language=ja}}</ref><ref>{{Cite web|和書|title=松本の松枯れを考える住民の会|url=https://www.facebook.com/MATSUMOTONONOYAKUSANPUWOKANGAERUJUMINNOKAI/posts/2096200610626305|website=www.facebook.com|accessdate=2019-05-26|language=ja}}</ref><ref>{{Cite web|和書|title=ネオニコチノイド系農薬等に関する質問主意書|url=https://www.shugiin.go.jp/Internet/itdb_shitsumon.nsf/html/shitsumon/a198021.htm|website=www.shugiin.go.jp|accessdate=2019-05-26}}</ref> |
|||
=== 批判への反論 === |
=== 批判への反論 === |
||
| 132行目: | 210行目: | ||
== 歴史 == |
== 歴史 == |
||
マツ枯れによると考えられるマツの集団枯損の記録は、1905年に長崎市周辺で発生したものにまで遡る。1914-15年頃には兵庫県赤穂市でも被害が発生し、以後九州山陽地方に被害が広がった。戦中戦後はマツ枯れ防除の余裕もなく、被害が拡大した。戦後は[[連合国軍最高司令官総司令部|GHQ]]の勧告のもと林野庁や各自治体が強力に伐倒駆除を進めた結果、一時被害は沈静化された。当時は家庭用燃料の3/4以上を薪が占めるなど燃料としての需要が旺盛だったため、マツ枯れ被害材も燃料用に飛ぶように売れたという。さらに[[里山]]アカマツ林では落枝落葉や球果([[松かさ]]、松ぼっくり)も肥料や燃料として利用されたため、林内にはあまり有機物が堆積せず貧栄養に保たれ、先駆樹種であるアカマツに適した環境が維持されていた。 |
マツ枯れによると考えられるマツの集団枯損の記録は、1905年に長崎市周辺で発生したものにまで遡る<ref>矢野宗幹(1913)長崎県下松樹枯死原因調査. 山林公報4</ref>。1914-15年頃には兵庫県赤穂市でも被害が発生し、以後九州山陽地方に被害が広がった。戦中戦後はマツ枯れ防除の余裕もなく、被害が拡大した。戦後は[[連合国軍最高司令官総司令部|GHQ]]の勧告のもと林野庁や各自治体が強力に伐倒駆除を進めた結果、一時被害は沈静化された。当時は家庭用燃料の3/4以上を薪が占めるなど燃料としての需要が旺盛だったため、マツ枯れ被害材も燃料用に飛ぶように売れたという。さらに[[里山]]アカマツ林では落枝落葉や球果([[松かさ]]、松ぼっくり)も肥料や燃料として利用されたため、林内にはあまり有機物が堆積せず貧栄養に保たれ、先駆樹種であるアカマツに適した環境が維持されていた。 |
||
しかし1950年代の[[燃料革命]]や化学肥料の普及によって、肥料や燃料としてのマツ林の落枝落葉の需要が減少して林内に放置されたため、土壌が富栄養化してアカマツには不適当な環境になり始めた。[[マツタケ]]の大幅減産もこの頃(1960年代前半)である。また、マツ材は強度が高いため屋根を支える梁などの建築用材としても生産・利用されてきたが、素材生産量は1957年の1551万立米をピークに減少に転じ、さらに1960年の木材輸入自由化後は価格競争にさらされ、1975年にはほぼ1/4にまで減少し、2004年にはわずか82万立米にまで落ち込んだ。このようなマツを巡る環境の下、防除の努力にもかかわらずマツ枯れ被害は1970年代から再び増加に転じ、1979年には被害材積が243万立米を記録している。以後毎年100万立米前後の被害が続いてきた。現在は保安林や景勝地などの保全と被害の北上を阻止することに重点をおいた防除が行われている。 |
しかし1950年代の[[燃料革命]]や化学肥料の普及によって、肥料や燃料としてのマツ林の落枝落葉の需要が減少して林内に放置されたため、土壌が富栄養化してアカマツには不適当な環境になり始めた。[[マツタケ]]の大幅減産もこの頃(1960年代前半)である。また、マツ材は強度が高いため屋根を支える梁などの建築用材としても生産・利用されてきたが、素材生産量は1957年の1551万立米をピークに減少に転じ、さらに1960年の木材輸入自由化後は価格競争にさらされ、1975年にはほぼ1/4にまで減少し、2004年にはわずか82万立米にまで落ち込んだ。このようなマツを巡る環境の下、防除の努力にもかかわらずマツ枯れ被害は1970年代から再び増加に転じ、1979年には被害材積が243万立米を記録している。以後毎年100万立米前後の被害が続いてきた。現在は保安林や景勝地などの保全と被害の北上を阻止することに重点をおいた防除が行われている。2015年現在、被害の北端は[[東北地方]]北部に達しており、日本海側が青森県南西部の[[深浦町]]付近、太平洋側はやや進行が遅く岩手県の中北部の[[八幡平市]]・[[滝沢市]]付近、[[三陸海岸]]沿いはさらに進行が遅く[[大船渡市]]付近となっている。 |
||
== 世界的な広がり == |
== 世界的な広がり == |
||
日本に近い東アジア諸国でもマツ枯れが問題になっている。中国では1982年に南京郊外で発生が確認され、[[バビショウ]](タイワンアカマツ)や日本産クロマツに被害が発生している。台湾では1970年代からリュウキュウマツのマツ枯れが報告されている。香港や韓国でも発生している。 |
日本に近い東アジア諸国でもマツ枯れが問題になっている。中国では1982年に南京郊外で発生が確認され、[[バビショウ]](タイワンアカマツ)や日本産クロマツに被害が発生している。台湾では1970年代からリュウキュウマツのマツ枯れが報告されている。香港や韓国<ref>S. C. Shin, (2008) Pine wilt disease in Korea. Pine Wilt Disease 26-32.</ref>でも発生している。韓国では日本産アカマツやクロマツの被害が多かったが最近ではチョウセンゴヨウの枯死も確認され始めたという<ref>H. Han, Y.-J. Chung, and S.-C. Shin,(2008) First report of pine wilt disease on ''Pinus koraiensis'' in Korea. Plant Disease 92(8): 1251-1251.</ref>。 |
||
ヨーロッパでは広く分布する[[ヨーロッパアカマツ]] ''Pinus sylvestris'' や[[フランスカイガンショウ]] ''P. pinaster'' が感受性であることから、ヨーロッパ各国が北米からのマツ材の輸入を禁止する措置をとって[[貿易摩擦]]にまでなった。1999年にはポルトガルのリスボン近郊でマツ枯れの発生が確認され、現在その封じ込めと根絶のための努力が行われている。 |
ヨーロッパでは広く分布する[[ヨーロッパアカマツ]] ''Pinus sylvestris'' や[[フランスカイガンショウ]] ''P. pinaster'' が感受性であることから、ヨーロッパ各国が北米からのマツ材の輸入を禁止する措置をとって[[貿易摩擦]]にまでなった。1999年にはポルトガルのリスボン近郊でマツ枯れの発生が確認され<ref> M. M. Mota; H. Braasch; M. A. Bravo; A. C. Penas; W. Burgermeister; K. Metge; E. Sousa,(1999)First report of ''Bursaphelenchus xylophilus'' in Portugal and in Europe. Nematology 1(7): 727-734.</ref>、現在その封じ込めと根絶のための努力が行われている。 |
||
== 脚注 == |
== 脚注 == |
||
| 146行目: | 224行目: | ||
== 参考文献 == |
== 参考文献 == |
||
* マツ枯れは森の感染症 森林微生物相互関係論ノート 二井一禎 (2003) 文一総合出版 |
* マツ枯れは森の感染症 森林微生物相互関係論ノート 二井一禎 (2003) 文一総合出版 ISBN 4-8299-2183-8 |
||
* 森林微生物生態学 二井一禎・肘井直樹 編著 (2000) 4.5 微生物と線虫を利用する昆虫の繁殖戦略―マツノマダラカミキリによるマツノザイセンチュウの伝播― 前原紀敏 朝倉書店 ISBN 4-254-47031-2 |
* 森林微生物生態学 二井一禎・肘井直樹 編著 (2000) 4.5 微生物と線虫を利用する昆虫の繁殖戦略―マツノマダラカミキリによるマツノザイセンチュウの伝播― 前原紀敏 朝倉書店 ISBN 4-254-47031-2 |
||
* [[きのこ]]の自然誌(新装版) 小川真 (1983) 築地書館 ISBN 978-4-8067-2198-7 |
* [[きのこ]]の自然誌(新装版) 小川真 (1983) 築地書館 ISBN 978-4-8067-2198-7 |
||
| 156行目: | 234行目: | ||
* [[五葉マツ類発疹さび病]] - アジア産の病原菌がアメリカのマツに壊滅的被害を与えている病気。 |
* [[五葉マツ類発疹さび病]] - アジア産の病原菌がアメリカのマツに壊滅的被害を与えている病気。 |
||
* [[クリ胴枯病]] - アジア産の病原菌が欧米、特にアメリカのクリに壊滅的被害を与えている病気。 |
* [[クリ胴枯病]] - アジア産の病原菌が欧米、特にアメリカのクリに壊滅的被害を与えている病気。 |
||
* [[ニレ立枯病]] |
* [[ニレ立枯病]](オランダニレ病) - アジア産の病原菌が欧米のニレに壊滅的被害を与えている病気。道管の閉塞による通水障害と病原体を昆虫が媒介するという点は似ている。 |
||
* |
* '''マツ材線虫病''' - 本項で解説。 |
||
=== マツ・針葉樹の重大な病気 === |
|||
* 五葉マツ類発疹さび病 - アメリカで五葉マツ類を植える時最も障害になる病気。上述。 |
|||
* '''マツ材線虫病''' - 日本でマツを植える時最も障害になる病気。本項で解説 |
|||
* スギ赤枯病 - スギ苗木を枯死に至らせる重要病害で成木にも感染し[[杉の溝腐病|スギ溝腐病]]を引き起こし木材の経済的価値を落とす。病原菌はアメリカ原産とされる。 |
|||
* 根腐れ - 原因菌は複数知られるが特に[[ナラタケ]]類、[[マツノネクチタケ]]類(特に欧米産)、[[エキビョウキン]]類などによるものが成木でも枯死被害が大きく有名 |
|||
* 心材部の腐朽 - 外見は正常だが木材の強度が下がり倒れやすくなることで間接的な死因になる。原因菌は複数知られる。 |
|||
=== 穿孔性昆虫やその共生生物による樹木の主な病気 === |
|||
* ニレ立枯病(オランダニレ病) - キクイムシが病原菌を媒介 |
|||
* [[ナラ枯れ]] - ナガキクイムシが病原菌を媒介 |
|||
* 北米マツ属樹木に対するに対する''Dendroctonus''属キクイムシの被害 |
|||
* [[モミ属]]・[[カラマツ属]]樹木に対するヤツバキクイムシ類の被害 |
|||
* [[ヤシ]]の赤輪病 - ゾウムシが病原線虫を媒介 |
|||
* オセアニア・北米のマツ属樹木に対する[[ノクチリオキバチ]]による被害 |
|||
* アメリカの[[トネリコ属]]樹木に対する[[アオナガタマムシ]]による被害 |
|||
=== 生物 === |
=== 生物 === |
||
* [[マツ属]]- 形態、生態、世界のマツ属植物一覧 |
* [[マツ属]]- 形態、生態、世界のマツ属植物一覧 |
||
* [[カミキリムシ]] |
* [[カミキリムシ]] |
||
** ヒゲナガカミキリ属 |
|||
* [[キクイムシ]] |
|||
:* [[カシノナガキクイムシ]] – ナラ類の病気を媒介する昆虫 |
|||
* [[ノクチリオキバチ]] – オセアニア等でマツ類の病気を媒介する昆虫 |
|||
=== マツと環境 === |
=== マツと環境 === |
||
* [[里山]] |
* [[里山]] |
||
| 170行目: | 263行目: | ||
* [[防風林]] |
* [[防風林]] |
||
== 外部リンク == |
== 外部リンク == |
||
=== 関連生物・病気 === |
|||
* [http://ss.ffpri.affrc.go.jp/labs/seibut/bcg/bcg00154.html 森林生物 マツノザイセンチュウ] |
|||
* [http://ss.ffpri.affrc.go.jp/labs/seibut/bcg/bcg00154.html 森林生物00154 マツノザイセンチュウ] 森林総合研究所資料 |
|||
* [https://www.ffpri.affrc.go.jp/labs/seibut/bcg/bcg00155.html 森林生物00155 マツノマダラカミキリ] |
|||
* [https://www.ffpri.affrc.go.jp/labs/seibut/bcg/bcg00253.html 森林生物00253 カラフトヒゲナガカミキリ] |
|||
* [http://www.ffpri.affrc.go.jp/labs/seibut/bcg/bcg00264.html 森林生物00264 オオコクヌスト] カミキリの天敵として期待される昆虫 |
|||
* [https://www.ffpri.affrc.go.jp/labs/seibut/bcg/bcg00284.html 森林生物00284 サビマダラオオホソカタムシ] 同上 |
|||
* [http://www2.kobe-u.ac.jp/~kurodak/Pine_wilt.html マツ枯れ…マツ材線虫病] 神戸大学森林資源学研究室 |
|||
* [http://www.apsnet.org/edcenter/intropp/lessons/nematodes/pages/pinewilt.aspx Pine wilt disease ― American Phytopathological Society] |
* [http://www.apsnet.org/edcenter/intropp/lessons/nematodes/pages/pinewilt.aspx Pine wilt disease ― American Phytopathological Society] |
||
* [http://www.na.fs.fed.us/spfo/pubs/howtos/ht_pinewilt/pinewilt.htm How to Identify and Manage Pine Wilt Disease and Treat Wood Products Infested by the Pinewood Nematodes] |
* [http://www.na.fs.fed.us/spfo/pubs/howtos/ht_pinewilt/pinewilt.htm How to Identify and Manage Pine Wilt Disease and Treat Wood Products Infested by the Pinewood Nematodes] |
||
* [http://entnemdept.ufl.edu/creatures/nematode/red_ring_nematode.htm red ring nematode - ''Bursaphelenchus cocophilus'' Cobb] 近縁線虫が引き起こすヤシの赤輪病についてフロリダ大学 |
|||
=== 技術 === |
|||
* [http://www.naro.affrc.go.jp/project/results/laboratory/nkk/1998/nkk98-19.html アコースティックエミッションによるトマトの水ストレスの検出] 農研機構HPによるAE法の応用事例紹介 |
|||
=== 薬剤 === |
|||
* [https://www.kankyo.metro.tokyo.jp/chemical/chemical/kids/insecticide/index.html 殺虫剤樹木散布編] 東京都環境局 化学物質の子どもガイドライン |
|||
=== 法律 === |
|||
* [https://laws.e-gov.go.jp/law/325AC0000000053 森林病害虫等防除法(昭和二十五年法律第五十三号)].e-Gov法令検索. 総務省行政管理局 <!-- 平成二十八年五月二十日公布(平成二十八年法律第四十四号)改正、2017年4月1日施行版を閲覧。--> |
|||
<!-- * [https://web.archive.org/web/20161011180158/http://law.e-gov.go.jp/haishi/S52HO018.html 松くい虫被害対策特別措置法] 電子政府の総合窓口e-gov -松くい虫被害対策特別措置法は1997年(平成9年)の特別措置法の失効に伴い、森林病害虫等防除法が改正され必要なものについては、この法律に取り込まれたので、2020年1月15日コメントアウトした。--> |
|||
{{林業}} |
|||
{{Normdaten}} |
|||
{{デフォルトソート:まつさいせんちゆうひよう}} |
{{デフォルトソート:まつさいせんちゆうひよう}} |
||
[[Category: |
[[Category:木の病気]] |
||
[[Category:マツ科]] |
[[Category:マツ科]] |
||
2024年12月20日 (金) 15:41時点における最新版
マツ材線虫病(マツざいせんちゅうびょう、英名:pine wilt disease)とは、マツ属(学名:Pinus)を中心としたマツ科樹木に発生する線虫の感染症である。病原体は北米原産で日本を含むアジアやヨーロッパのマツ類に枯死を伴う激害をもたらしている。日本における病気の汚染地域は徐々に拡大しており、2010年以降、本州以南の46都府県全てで確認されている。関係者の間ではマツ枯れと呼ばれることが多い。本項でも「マツ枯れ」を用いる。行政用語としては松くい虫が用いられる。
世界三大樹木病害(ニレ立枯病、クリ胴枯病、五葉マツ類発疹さび病)に本病を加えて世界四大樹木病害と呼ぶことがある。また、外見上類似した病害としてナラ枯れ(ブナ科樹木萎凋病)がある。
症状と診断
[編集]典型的な症状としては盛夏から秋にかけてそれまで正常であったマツの針葉が急速に色あせて最終的には褐変する。針葉の褐変は症状の最終段階であり、それに先だって外見は正常なまま樹脂(いわゆる松脂)の滲出が減少する。健康なマツは幹に傷を付けると大量の樹脂を傷口に分泌するが、この病気を発病したマツは樹脂の量が著しく減少し全く出さないことも多い。このため、早期の診断には幹にピンを刺したりポンチで穿孔したりして樹脂滲出異常の有無を調べる[1][2]。この簡易判定方法は発見者の小田久五に因み「小田式判定法」などと呼ばれる。また、発病した個体の幹には多数の穴が見られることが多い。これはこの病気に限ったことではないが、マツが弱ってくるとキクイムシやカミキリムシが集まってくるためである。
条件によっては典型的な経過とならず、樹脂滲出が止まっても外見が正常なまま翌年まで生存することがある。冷涼な地方ではこのような経過をたどる個体が温暖な地方より多い。それらの個体は翌年の春から初夏に枯死して「年越し枯れ」と呼ばれたり、さらに遅れて通常のマツ枯れシーズンに至って枯れて「潜在感染木」と呼ばれることもある。
-
 枯死したヨーロッパクロマツP. nigra
枯死したヨーロッパクロマツP. nigra -
 枯死したリュウキュウマツ(P. luchuensis)
枯死したリュウキュウマツ(P. luchuensis) -
 葉が褐色に変色したヨーロッパクロマツ(P. nigra)
葉が褐色に変色したヨーロッパクロマツ(P. nigra)



原因
[編集]マツノザイセンチュウ Bursaphelenchus xylophilus (Steiner & Buhrer) Nickleと呼ばれる線虫の一種の感染による。以下、特に断りのない限り線虫と略す。かつてはマツ枯れの原因は昆虫ではないかという説が長くあり、「松くい虫」などの病名にも名残が見られる。この説では「松くい虫」として何種類かのキクイムシやゾウムシ、カミキリムシなどが挙げられており防除方法の研究が行われていた[3]。確かに枯死木には多数の昆虫が見られるが、これらはマツが弱ってから侵入したのであって因果が逆であることが分かっている。また、枯死木を伐採するとその切り口が青色に変色していることが多いことから、これを引き起こす青変菌がマツ枯れの原因ではないかという説もあった。1969年、林業試験場(当時、現:森林総合研究所)九州支所の職員だった清原友也と徳重陽山の2人によって九州のマツ枯れ枯死木から日本では見慣れない線虫が非常に高い確率で発見される現象が報告され[4]、さらにその後この線虫を接種したマツが枯死したことから線虫がマツを枯死させる原因であることが判明[5]。この線虫は線虫学者の真宮靖治らによって新種Bursaphelenchus lignicolousとして記載され[6]、和名はマツノザイセンチュウとされた[7]。
線虫の感染からマツの枯死に至るメカニズムは完全には解明されていないが、最終的には樹体内部の管類の閉塞、特に仮道管の閉塞が広範囲で起こり水を吸い上げられなくなって枯死することが分かっている。抵抗性種では線虫の移動が抑制され管類の閉塞が部分的であり、致命的な全身の通水障害を起こさないために枯死には至らないという。共に抵抗性だが分類学的には遠縁のストローブマツ(Pinus strobus)とテーダマツ(P. taeda)を観察し比較した結果では、抵抗性種であっても種類によってその防御機構が異なっていることを示唆するような報告がある[8]。
近年、この閉塞の原因は線虫や細胞による物理的な管の詰まりではなく、キャビテーションという現象によって細い管内に気泡が発生することにより管内部に空洞が形成され、この空洞が栓の役割を果たして樹液の流れを妨げること(ガスエンボリズム=気体塞栓症)が継続的に発生していることが明らかになってきた[9][10]。キャビテーションによる仮道管の閉塞自体は健全個体でも乾燥時等に見られるが一時的であれば可逆的である。なぜ継続的なキャビテーションが発生してしまうのかは分かっていない。線虫もしくは線虫に抵抗したマツが作り出す毒素説、菌類などの第三者が関わる説、樹液が変性し表面張力を失う説などがある。
キャビテーション・エンボリズムの診断方法として着色溶液をマツに吸わせてから切断して断面を観察するという古典的方法の他にも、水が途切れる際に発生する音の一種、アコースティック・エミッション(Acoustic Emission, AE)を観測するという非破壊的方法が提案・実用化されており[11]、蒸散速度や光合成速度の低下を観察するという従来からの非破壊的方法に加えてこのような面からも樹木の水分異常を観測できるようになってきている。
線虫は病原性を持ちマツを枯らすことが出来るが、他のマツへの移動手段を持たない。これを助けるのがヒゲナガカミキリ属(Monochamus)のカミキリムシである。線虫は蛹室内にいるカミキリムシ新成虫の気門に侵入し、脱出したカミキリムシと共に他のマツへと移る。
病原線虫 Bursaphelenchus xylophilus
[編集]この線虫が初めて見つかったのはアメリカ・ルイジアナ州で、現地原産のダイオウマツ(Pinus palustris)木材中から発見され、当初はAphelenchoides xylophilusとして記載された[12]が、その後Bursaphelenchus属に移された。日本で見つかりマツへの病原性を示す線虫は前述のようにBursaphelenchus lignicolousとして新種記載されたが、これも後に移動され同一種扱いとなっている。
B. xylophilusは体長1mmほどの三日月形で雌雄の区別がある。餌はマツの柔細胞と菌類であり、セルロース分解酵素(セルラーゼ)を分泌してマツの細胞を破壊・摂取する。実験室内では菌類だけを餌に培養することが可能である。実験室内での培養温度は餌にする菌類の種類にもよるが15℃以上で増殖、25℃程度で活発に増殖するという[13]。ただし、何世代にもわたって実験室内で培養し続けると病原性と増殖力が著しく低下してしまうという報告がある[14]。野外においても病原性の異なる系統が共存しているという[15]。病原性の違いの一つにセルロース分解酵素の強弱の差があるとされる[16]。
カミキリムシ(以下カミキリ)の蛹の気門に入った線虫はカミキリ成虫が羽化するまで待つ。カミキリが羽化し、成虫が餌として齧る健康なマツの若枝から健康なマツの樹体内に侵入する[17]。この時期は後述のカミキリの生活環にもよるが、一般に5―8月の初夏~盛夏の時期である。マツ体内では樹脂道を使い移動[18]、交尾・産卵を繰り返しながら増殖する。このとき抵抗性の種では線虫の移動は部分的であるが、感受性の種は全身に移動し分布するようになる。また、線虫の増殖は感受性種であっても最初は穏やか、マツが目に見えて弱り始めてから爆発的に増えると言う。マツは線虫の侵入(感染)から2週間程度で樹脂の分泌が減少を始め、2-3カ月程度で急激な葉の変色と枯死に至る。弱ったマツはカミキリの格好の産卵場所であり、カミキリが産卵に来る。線虫は枯死木の中で青変菌などを食べながら増殖し[19][20]カミキリに乗り移る機会を窺う。年が明けカミキリが蛹から羽化して成虫になる際に線虫はカミキリ気門に潜り込みカミキリと共に新しいマツへと移る。
-
 雄線虫
雄線虫 -
 雄線虫と突起状の生殖器
雄線虫と突起状の生殖器 -
 線虫を採取するベールマン装置模式図
線虫を採取するベールマン装置模式図



ニセマツノザイセンチュウ Bursaphelenchus mucronatus
[編集]マツノザイセンチュウは北米原産の外来種とされるが、日本には在来種として同属のニセマツノザイセンチュウ B. mucronatus Mamiya et Enda が分布している。本種と比べると弱い病原性を持つ[21]が自らマツに病気を起こすことはできないため、すでに何らかの理由で衰弱していたマツを生息場所にしている[22]。マツノマダラカミキリも同様で、衰弱して樹脂などによる防御力の弱まったマツに産卵するのが本来の生活様式であった。ニセマツノザイセンチュウもマツノマダラカミキリを媒介者とするが、マツノザイセンチュウとは異なりカミキリを離脱するのは後食時ではなく主に産卵時である。つまり、健全なマツを発病させて媒介者であるカミキリを呼ぶ代わりに、衰弱したマツに媒介者と同時に侵入していると考えられている。ニセマツノザイセンチュウとマツノマダラカミキリは、マツ林に時折生ずる被圧木や折損枝、風倒木などの衰弱木や新鮮枯死材を利用して、細々と生活していた生物であると考えられている。
その他の類縁線虫
[編集]線虫という生き物は非常に多種類が知られ、土壌中で腐植を食べるような物から昆虫や動物に寄生して病気を引き起こすものまでいろいろである。Burusaphelenchus属線虫は生態面で植物寄生性であり、媒介を昆虫に頼るのを特徴としている。現在100種以上が知られており、大きく3グループに分けられるという[23]。日本においてはマツノザイセンチュウ、ニセマツノザイセンチュウの他にも、ウコギ科樹木に付くタラノザイセンチュウ (Burusaphelenchus luxuriosae)、クワに付くクワノザイセンチュウ (B.conicaudatus)などが知られる。宿主に対する病原性は一般的に低く、何らかの原因で衰弱していたものを枯死に追い込むことが出来る程度のものが多いとされる。マツ属枯れ木で見つかるB. douiなど病原性は無いのではないかとも言われる種もある。しかしながら、中には高い病原性を持つものも存在し、特に有名なのが本病のB. xylophilusと中南米やカリブ海諸国のヤシ類プランテーションで赤輪病(英名: red ring disease)を引き起こし大きな被害を出しているB. cocophilusの2種。ただし、これら2種共に被害を出しているのは土着の植物ではなく持ち込まれた植物に対してである。最近では他の種でも病原性が比較的強いものも報告されており、マツ類苗木に病原性を示したB. sexdentatiなどがある[24]。
媒介昆虫
[編集]マツノザイセンチュウが発見されたのに続き、この線虫がマツノマダラカミキリ(Monochamus alternatus)というカミキリムシによって媒介されて感染拡大を引き起こしていることが発見報告された[17]。このカミキリムシはマツ枯れ被害木からよく見つかることから以前から「松くい虫」候補の一つとして名前が挙がっていた昆虫である。
マツノマダラカミキリはヒゲナガカミキリ属(Monochamus)に属する小型のカミキリムシで、日本ではこの種以外にも近縁種が複数分布する。マツ枯れ被害にはマツノマダラカミキリ(M. alternatus)の他に同属のカラフトヒゲナガカミキリ(M. saltuaris)も関与していることが報告されているが、後者は前者に比べて線虫の保持数は著しく低く、関与は部分的と言われる。属名の通り両種とも長い触角(ひげ)を持つが、体の方は何れも1-3cm程度と小型。色もマツノマダラは茶色の斑(まだら)模様、カラフトヒゲナガは黒色と地味である。アメリカでの媒介は同属のM. carolinensisなど数種、スペインではM. galloprovincialis という種が主に関わっているという。マツノザイセンチュウの媒介では脇役のカラフトヒゲナガカミキリではあるが、ニセマツノザイセンチュウの媒介では主役であり主要な媒介昆虫であるとされる[25]。
これらのカミキリの成虫は特に弱った木や枯死したばかりの木に好んで産卵し、幼虫はマツ(正確にはマツ属以外のマツ科植物も食べる)の材を食べて育つ。産卵は晩春から夏にかけて行われる。幼虫は初め樹皮直下の組織を食べるが、冬になるとより内部に潜り込む。冬になるまでに一通りの成長を終えた幼虫はさなぎを経て、6月-7月にかけて羽化する。カラフトヒゲナガの場合はもう少し早いという[26]。成虫の餌はマツの若い枝であり、材から脱出するとこれを食べに向かう(昆虫学の用語では羽化後の摂食を後食と言う)。後食をしていくうちに性成熟したカミキリは繁殖相手を求め始め、やがて交尾・産卵し夏の終わりには姿を消す。
マツ枯れ被害が確認されていない北海道においてもシラフヨツボシヒゲナガカミキリ(M. urussovii)という近縁種が分布する。マツ属樹木に乏しい北海道の環境のためか、この種が餌として利用するのはマツ科トウヒ属(Picea)のエゾマツ(Picea jezoensis)やアカエゾマツ(Picea glehnii)、同科モミ属(Abies)のトドマツ(Abies sachalinensis)だという。このカミキリはこれら樹木の害虫として認識されているが、線虫が北海道に侵入した場合に線虫を媒介するのかどうかについてはよくわかっていない。ただし、この種はニセマツノザイセンチュウをエゾマツやトドマツに媒介することは確認されている[27]。
他のBursaphelenchus属線虫を媒介する昆虫にはカミキリムシ、キクイムシ、ゾウムシ、クワガタムシ、ある種のハナバチ等が知られるという[23]。なお、マツノザイセンチュウに関しては近年根の罹病木と健全木の癒合部分からも感染することが明らかになっている[28]。
-
 マツノマダラカミキリ(M. alternatus)
マツノマダラカミキリ(M. alternatus) -
 カラフトヒゲナガカミキリ(M. saltuarius)
カラフトヒゲナガカミキリ(M. saltuarius) -
 マツの枝に付く北米産近縁種M. mutator
マツの枝に付く北米産近縁種M. mutator -
 ヨーロッパ産近縁種M. galloprovincialis
ヨーロッパ産近縁種M. galloprovincialis




青変菌
[編集]青変菌(英名:blue stain fungi)は名前の通り木材を青く変色させることで知られる菌類でCeratocystis属、Grosmannia属、Ophiostoma属など複数の属に跨って知られる。マツ枯れによる枯死木を伐採すると切り口が青く変色していることが多く、かつては青変菌がマツ枯れの主原因ではないかとも言われていたこともある。日本では青変菌は木材の経済的価値を落とす菌として有名だが、世界的にみると欧米のニレ属樹木に壊滅的被害を出し続けているニレ立枯病(オランダニレ病)を引き起こすOphiostoma ulmiやアメリカのナラ類のoak wilt病(和名未定)を引き起こすCeratocystis fagacearum、アメリカ西部のキクイムシの一種(Dendroctonus ponderosae)が引き起こすコントルタマツ(Pinus contorta)の病気などのように病原性の強いものも複数知られる。しかしマツ枯れの場合マツが枯死してから青変菌が増殖することなどから主たる原因ではないとされた[29]。マツ枯れ材に青変菌が見られるのは主にカミキリムシやキクイムシが体に付けて持ち込むからだという。
-
 青変菌の繁殖した樹木の切断面、マツ枯れ枯死木もこのようになることが多い
青変菌の繁殖した樹木の切断面、マツ枯れ枯死木もこのようになることが多い -
 世界的に有名なニレ立枯病(オランダニレ病)
世界的に有名なニレ立枯病(オランダニレ病) -
 アメリカのoak wilt病(和名未定)
アメリカのoak wilt病(和名未定) -
 キクイムシと青変菌により枯死した大量のコントルタマツ. アメリカ・コロラド州
キクイムシと青変菌により枯死した大量のコントルタマツ. アメリカ・コロラド州




その他の原因説
[編集]線虫はコッホの原則を満たし大規模なマツ枯れを引き起こしている原因であることはほぼ間違いないが、原因・主因は他にあるとする見方も少ないながら存在する。線虫以外の病原体説、大気汚染説、酸性雨説、土壌の富栄養化によるマツの根の衰弱説などが特に有名。これらの要因でマツが衰弱し、時として枯死に至ることは確かにあり[30][31][32]、実際の山林のマツ枯れ被害には線虫を主因にこれらの要因を加えた複合的な問題が存在すると考えられている。



発病に関する因子
[編集]マツの種類と感受性
[編集]マツの種類により線虫への感受性に差がある。日本産種は何れも感受性が高く、特にクロマツ(Pinus thunbergii)やチョウセンゴヨウ(P. koraiensis)は弱い。線虫が天然分布する北米産種は特に東部に分布する種を中心に線虫に抵抗性を示すものが多い。興味深いことにアジア産種であっても抵抗性を示す種がいくつか知られる、逆に北米種であっても西部やメキシコに分布するものを中心に弱いものもある。以下、古野(1982)[33]における野外での観察による結果を基に、二井・古野(1979)[34]、や清原・徳重(1971)[5]も加えて感受性についてのリストを示す。人工的な線虫接種試験とカミキリの嗜好が加わる野外観察では結果が変わることが予想される。また、線虫接種試験では接種時の樹齢や接種量などにより差があり、抵抗性種であっても苗木に接種したり、過剰に接種すると枯れる場合がある。ゆえに抵抗性の強弱は研究者によって多少の相違があるので参考程度にされたい。
種名の後はその種の主な分布地を示す、米東=アメリカ大陸東部、米西=アメリカ大陸西部・メキシコ、亜=アジア、欧=ヨーロッパ
- 強抵抗性
- テーダマツ(Pinus taeda、米東)、リギダマツ(P. rigida、米東)、ダイオウマツ(P. palustris、米東)、スラッシュマツ(P. elliotti、米東)、ニイタカアカマツ(P. taiwanensis、亜)
- やや抵抗性
- ストローブマツ(P. strobus、米東)、バンクスマツ(P. banksiana、米東)、コントルタマツ(P. contorta、米西)、バビショウ(タイワンアカマツ, P. massoniana、亜)、レジノーサマツ(P. resinosa、米東)、クロマツ×バビショウ雑種第一代(F1)、P. tabuliformis(亜)
- やや感受性
- シロマツ(P. bungeana、亜)、モンチコラマツ(P. monticola、米西)、P. storobiformis(米西)、ゴヨウマツ(P. parviflora、亜)、アカマツ(P. densiflora、亜)、フランスカイガンショウ(P. pinaster、欧)、ヨーロッパアカマツ(P. sylvestris、欧亜)、ポンデローサマツ(P. ponderosa、米西)、ラジアータマツ(P. radiata、米西)、
- 強感受性
- クロマツ(P. thunbergii、亜)、チョウセンゴヨウ(P. koraiensis、亜)、リュウキュウマツ(P. luchuensis、亜)、ヨーロッパクロマツ(P. nigra、欧)、モンタナマツ(P. mugo、欧)、P. muricata(米西)、P. leiophylla(米西)
このリストには載せてないがハイマツ(P. pumila)や屋久島・種子島に分布する絶滅危惧種ヤクタネゴヨウ(アマミゴヨウ、P. amamiana)[35][36]も感受性である。ただし、ハイマツなどは比較的寒冷地に分布するために実験室以外の野外で影響を受けることは今のところあまりないとされる。
また、マツ属以外のマツ科樹木のうちモミ属(Abies)、トウヒ属(Picea)[37]、カラマツ属(Larix)、ヒマラヤスギ属(Cydrus)の一部樹木についてマツノザイセンチュウによる枯死例が報告されている。
その他の要因
[編集]マツ枯れの主因は線虫だが、様々な要素が誘因となって発病を促進したり、また緩和要素となって発病を抑制したりする。
代表的な誘因は高温と乾燥によりもたらされる水ストレスである。マツ枯れは萎凋病であり、最終的には材の通導阻害によって枯死に至るため、水ストレスは発病を促進する。逆に1993年のように夏が低温で雨が多いと、これが緩和要素となって枯死にまでは至らない個体が増加する。
大気汚染や酸性降下物など環境汚染も誘因となる。これらは単独ではストレスとはなっても、健全なマツを枯死にまで追い込むことはあまりないと考えられている。しかし線虫に感染したマツでは、これらの影響により通常であれば発病しない条件でも枯損が生じることがある。
マツ林の放棄に伴う土壌の富栄養化も間接的誘因となることが以前より示唆されている。健全なマツは吸収根のほとんどが外菌根となっており、それにより養分・水分の乏しい環境に適応しているとされているが、土壌の富栄養化によって吸収根は土壌表層に集まり、なおかつ菌根の発達が悪くなる。土壌の表層は夏の乾燥の影響を受けやすい部分でもあり、このような状態では水ストレスに敏感になると考えられている。
松枯れの蔓延とその影響
[編集]生態面
[編集]マツ枯れの蔓延によってクロマツ林が壊滅し、広葉樹が侵入してきた海岸の事例では上層の構成種の変化だけでなく、下層の植生や土壌成分に変化があったことが報告されている。[38]。
人間生活
[編集]日本における防除方法とそれに対する批判
[編集]方法
[編集]現在、材線虫病の対策は薬剤の事前投与による発病予防と枯死木およびカミキリの駆除による病気の拡散防止が行われている。発病後の有効な対策は見つかっていない。
カミキリムシの駆除
[編集]線虫を媒介するカミキリムシを駆除し材線虫病の拡大を防ぐ方法、幼虫・成虫それぞれに方法がある。
幼虫を標的としたものとしては、カミキリが衰弱状態や枯死したばかりのマツに好んで産卵する性質を利用し、秋から翌年春にかけて枯死したばかりの木を伐採する方法がある。その後伐採木の焼却、粉砕、もしくは薬剤散布によって材の中のカミキリ幼虫を殺す。なお、薬剤散布の場合は幼虫が材内部に食い進む冬以前に行わないと殺虫効率が悪いという。これらは「伐倒駆除」と呼ばれ重労働である。近年薬剤散布方法が見直され、被害木を適当な大きさに玉切りしたあと薬剤散布とビニールシートでの被覆をセットで行うようになってきており、単なる散布と言うより燻蒸に近くなった。この方法の利点として薬剤の飛散がなく殺虫効率も高いこと、さらに破砕や焼却といった作業に比べてその場で処理できることが挙げられる。薬剤には有機リン化合物やカルバミン酸系化合物(通称カーバム剤・カーバメート剤、商品名NCS等)などが使われる。
成虫を標的にしたものとしては成虫の羽化してくる初夏(5-7月)に、カミキリが後食するマツの若い枝がある樹冠部に殺虫剤を散布し、カミキリを殺すことで線虫の感染を防ぐというものである。後食の開始と線虫がカミキリから脱出しマツに感染するのには僅かではあるがタイムラグがあるので[17]この間にカミキリを殺すことは感染防止に有効とされる。主に大型噴霧器による地上からの散布かヘリコプターによる空中散布が採られる。薬剤には後述のように農薬の定番有機リン系か、最近はそれに比べて人畜毒性の低いと言われるネオニコチノイド系が使われる。散布には周辺のドリフト被害などからの批判もある。また、農薬の種類によっては森林認証制度を利用できなくなる場合がある。最近では薬の他に昆虫病原糸状菌、特にその中でもBeauveria属菌を使う殺虫方法なども提案されている。ただし、この病原菌も農薬同様非選択性的性格が強く様々な昆虫を殺してしまう。
カミキリ特に幼虫時期を対象とした天敵動物による防除、いわゆる生物農薬についても研究が行われているが、広く実用化されたものは無い。鳥類ではキツツキの一種のアカゲラ[39]、寄生性・捕食性昆虫としてホソカタムシ類[40][41][42]、コクヌスト類[43]、寄生蜂[44]等が注目されており、生息密度を上げることや効率的な捕食・寄生を行う方法などが研究されている。線虫の移動を助けるカミキリではあるが、ある種の線虫はカミキリに寄生する。マツノマダラカミキリから見つかった線虫はContortylenchus genitalicolaとして新種記載され[45]、しかも卵巣に寄生するということから不妊化効果が期待されたが、完全な不妊雌成虫を作り出すのは難しくこの計画はとん挫した。なお、海外の事例ではオセアニアにおける北米原産マツ類に枯死被害をもたらす侵入昆虫ノクチリオキバチ(Sirex noctilio)においてキバチ雌成虫を不妊化する寄生線虫が見つかっており[46]、この線虫を用いた被害のコントロールも実現している[47]。また、キバチの線虫とは属が異なるが、日本においてもスズメバチ女王蜂を不妊化する寄生線虫が見つかっている[48]。
寄生線虫に代わって注目されているものに、ボルバキアがある。ボルバキアは昆虫や線虫に寄生する細菌で宿主雌成虫の産卵により次世代へと繋がるために、宿主昆虫の生殖機構を乗っ取り宿主雌個体、ひいては自分に有利に利用することが多い。他にも犬のフィラリア症ではフィラリアに感染したボルバキアが病原性を高めたり、ある系統のボルバキアが感染したネッタイシマカはデング熱を媒介しにくくなる、宿主昆虫の寿命を縮めるなどの面白い性質が知られるものもある。近年マツノマダラカミキリを調べた結果、ボルバキアは単なる感染ではなくカミキリの常染色体に組み込まれているといい、遺伝子の水平伝播を起こしたことが示された[49]。昆虫のような高等生物でこのような水平伝播は珍しいという。ボルバキアは前述のように宿主の生殖機構のコントロール能力を持つことがあるためカミキリについても不妊虫放飼のような効果があるのではないかという仮説の基で研究が進められている。
他にはマツが弱った時に発する匂いを模したもので、産卵期のカミキリを誘因する薬剤などもある。マツノマダラカミキリでの誘因調査事例では比較的上の方によく集まるという[50]。マツノマダラカミキリ成虫が忌避行動をとる周波数の音波があることが発見され、農薬に変わる新たな防除手段になるのではないかと期待されている[51]
特定の林分でマツ枯れを防除する場合、侵入初期に異常を示した少数のマツを的確に伐倒処理すれば、林内で感染サイクルが回って被害が拡大するのを防ぐことができる。しかし近隣に激害状態の林分があれば、そこから線虫保持カミキリが侵入することは防げない。理論上は林内でカミキリが発生して激害とならないよう薬剤散布による殺虫、近隣林分の樹種転換によってカミキリの飛来が充分少なくなるまで持ちこたえることができれば、防除は成功すると考えられている。日本における完全な根絶例は沖永良部島の一例で1995年に根絶宣言がなされている。この事例では空中散布によるカミキリ駆除の効果が高かったことが指摘されている[52]。この他に鹿児島県の吹上浜[53]や佐賀県の虹ノ松原で被害をほぼ抑えることに成功している。ただし、吹上浜については2008年頃から薬剤散布が行われなくなって再び激害化してしまい、図らずも空中散布の有効性が示される結果となった。
発病後一気に枯死に至らず、年が明けた春先、場合によっては夏近くまで生存する個体があり寒冷地では半数以上がこのタイプと言われる。カミキリ成虫が羽化する時期にこれらの衰弱した個体がある場合、マツがカイロモンとなるエタノールやテルペン類を発して性成熟したカミキリを誘引するため、防除の障害となる。
-
 Beauveria bassianaは多犯性で様々な宿主に感染する。これはバッタだが、カミキリも同じ菌が殺す
Beauveria bassianaは多犯性で様々な宿主に感染する。これはバッタだが、カミキリも同じ菌が殺す -
 アカゲラ
アカゲラ

.jpg)
線虫の駆除
[編集]薬剤を樹幹に注入するか土壌に散布し根から吸収させる。効果では線虫増殖抑制作用のものと殺線虫作用のものに大別される。
代表的なものにモランテル(酒石酸塩が商品名グリーンガード)、メスルフェンホス(商品名ネマノーン)、レバミゾール(塩酸塩が商品名センチュリー等)、マクロライド系と呼ばれ、土壌中の細菌が産生する3種、ネマデクチン(商品名メガトップ)・エマメクチン(商品名ショットワンツー)・ミルベクチン(商品名マツガード)などがある。これらの薬剤の有効成分の多くは人を含む動物における線虫感染症にも用いられており、中には殺虫剤として使われるものもある。その他の薬でも効くものがあり、線虫も人や昆虫と同じくアセチルコリンを神経伝達物質として使うので神経伝達作用を阻害する作用の薬、例えばアセチルコリンの速やかな分解を阻害する有機リン剤なども線虫に有害である[54][55]。
マツ材線虫病は感染後急激に発症枯死に至り、また管類の閉塞が見られることもあって、投与は線虫の感染前に予防的に行われることが多い。防除効果は極めて高いがマツ1本あたり数千円の薬剤費用がかかり、かつ1-3年に一回の継続的な投与が必要だという費用的な問題の他、ドリルによって穴を開けることや傷口の不適切な処理による傷害、また薬害が出る場合もある等の欠点がある[56][57]。薬剤の成分調整や施用方法の改良によりこれらの問題は改善されつつあるが、費用面・労働面の負担が依然として農薬散布に比べ大きく広範囲に用いるのは現実的ではない。主に各地の銘木や貴重な個体群、個人所有の庭木などに対して用いられる。
カミキリの場合同様、天敵生物の利用もいくつか研究されているが実用に至ったものは無い。マツ丸太に対して線虫とヒラタケ等の線虫捕食菌を共に接種した結果では、マツノザイセンチュウの増殖を抑制することがしめされている[58]。またトリコデルマ(Trichoderma)属菌やアクレモニウム(Acremonium)属菌は線虫に対して有害であるとされる[19]。マツノザイセンチュウは系統によって病原性が大きく異なり、また強毒性の系統でも培養を重ねると病原性を失う[14]。この病原線虫の発見者の一人である清原友也は弱毒性系統の線虫を事前に接種しておくと、後に強毒性系統の線虫を接種した時に枯死しにくいというワクチンのような誘導抵抗性効果がある[59]という面白い現象を報告しているが、これも実用化には至っていない。
他の植物寄生線虫による被害、特に農作物における線虫の被害は土壌病害という扱いになるものが多く、薬剤燻蒸の他にも連作を避けたり、殺線虫効果のあるマリーゴールド等の植え付け、天地返しなどの伝統的対処法が知られている。細菌の一種バチルス・チューリンゲンシス(Bacillus thuringiensis)の産生する毒素は昆虫に有害、人に無害ということでBT剤などと呼ばれ農薬に用いられてたり、毒素を作り出す遺伝子を農作物に組み込んで遺伝子組み換え作物として利用されたりしている。近年この毒素が昆虫のみならずある種の線虫にも有害であるという報告がなされており[60]、農作物だけでなく他の分野への応用が期待される。
-
 薬剤の樹幹注入風景
薬剤の樹幹注入風景 -
 レバミゾール
レバミゾール -
 エマメクチン
エマメクチン -
 ミルベメクチン
ミルベメクチン -
 チオナジン
チオナジン
.jpg)



被害材の移動制限
[編集]在来、外来問わずマツ枯れ被害材の安易な移動は、木材内に線虫保持カミキリがいる可能性を考えると慎むべきである。離島での発生はもちろん、そもそも日本での発生も材内にいた線虫保持カミキリから始まったと考えられている。
日本では森林病害虫等防除法第3条第5項に基づき移動制限・移動禁止措置が義務付けられている[61]が、平成の時代に入っても奄美大島など離島部で新規にマツ枯れの報告があるところを見ると守られているとは言い難いのが現状である。日本国外でもMonochamunos属カミキリムシの侵入を警戒している国は多く、木材輸出入の際には剥皮や乾燥を求められることが多い。
予防的な在来マツの伐採、樹種の切り替え
[編集]激害林分及び線虫が侵入すれば激害となることが予想される林分は、積極的に伐採して樹種転換を図り、被害拡大を食い止めることも行われている。2006年には青森県においてマツ枯れの北上を阻止するために隣接する岩手・秋田の2県のうち、病気の北上の著しい秋田県との県境に約2km幅の防除帯を設けマツをすべて除去するという対策が採られた。なお、青森県内では県境から遠いにも拘らず蓬田村において2010年1月に線虫感染による枯死木が初確認され、久々に新規都道府県での感染確認となった。その後2013年には危惧されていた通り秋田県に隣接する深浦町で枯死木が確認されるなど徐々に被害が出始めている。
また、カミキリは線虫の感染に限らず弱ったマツを産卵対象として好むために、林内の競争に負けて弱った個体を間伐等で間引くのはカミキリを誘因・産卵させないために有効とされる。
抵抗性品種の植栽・開発
[編集]かつては抵抗性の北米種を植えることが行われた。テーダマツやリギダマツが代表的である。特にテーダマツは原産地では樹高40-50mに達することもある大型種で在来アカマツと比べて苗木の成長もきわめて速い[62]などと対マツ枯れ以外の面も期待されたが、地上部に比べて根の発達が貧弱で風で損傷しやすいことや、外来種問題などから現在は廃れてしまった。他にも在来のクロマツにバビショウ(タイワンアカマツ、P. massoniana)を掛け合わせた雑種などが開発されたことがある。バビショウはアジア産種でありながら線虫に対して強い抵抗性があり、この雑種も抵抗性を持ったという。ただし、これも線虫には抵抗性があっても他の害虫に弱いという欠点があり広い普及には至らなかった[63]。
現在主流なのは在来種の抵抗性個体の選抜である。マツ枯れ被害の大きな森林でも枯死せず生き残った個体から種子を採り、発芽させた苗木に線虫を投与して枯死しなかったものを選抜する。実生の子は親のクローンではないために必ずしも抵抗性を持つとは限らない[64]ために、線虫の接種による抵抗性の検定が欠かせず、手間と費用がかかるのが欠点である。挿し木などの無性繁殖では子供は親の形質をそのまま受け継ぐために、抵抗性系統の母樹から低コストで効率的に生産ができると考えられている。一方でマツは一般的に挿し木による繁殖が難しいことで知られており、効率的な発根・定着条件を探す研究が続けられている[65][66][67]、近年の研究では発根率の良い若い苗木時代に植物ホルモンのサイトカイニンを投与してやると発根率の良い挿し穂が量産できるとされている。アカマツには抵抗性系統が比較的多く見つかっているが、クロマツの抵抗性系統は少ない。また、これらについても抵抗性はあるが、樹形や成長速度などの面は考慮されておらず課題が残る。
病理的に不明な部分があること、日本人の遺伝子組み換え作物に対する嫌悪感もあってか遺伝子組み換えの抵抗性マツを作ろうという動きは殆どない。海外種との交雑実験も前述のように廃れてしまった。
防除事業への批判
[編集]現状の松くい虫防除事業には、一定の批判がある。
カミキリ成虫を標的にした農薬の空中散布は根強い反対意見がある。農薬散布は1977年(昭和52年)4月18日付法律第18号の松くい虫被害対策特別措置法 で制度化された。(その後、1997年(平成9年)の特別措置法の失効に伴い、森林病害虫等防除法が改正され必要なものについては、この法律に取り込まれている。同第七条の二、防除実施基準など)カミキリ駆除を目的として散布される農薬には大きく分けて有機リン系とネオニコチノイド系があり、何れも昆虫の神経に作用する。有機リン系化合物は現在の農薬の定番とも言える薬剤ではあるがサリンなどの化学兵器にもなる猛毒ガスも知られ、ネオニコチノイドはミツバチの異常行動の原因として疑われるなど何れも印象はあまり良くない。前者にはフェニトロチオン(商品名MEP剤、スミチオン、スミパイン)やマラチオン(商品名マラソン)、プロチオホス(商品名プロチオン、ソビーT)。後者にはアセタミプリド(商品名マツグリーン)やチアクロプリド(商品名エコワン、エコファイター)などがある。これらの農薬はカミキリ以外の昆虫にも有害(所謂「非選択性」の農薬)であるため、生物多様性などに悪影響を与えるのではと言う声もある。
有機リン・カーバメート(有機窒素)などのコリンエステラーゼ阻害剤による野生生物による影響としては以下に示すような報告がある。ただし、上に挙げたようなマツ枯れに実際に使う薬剤だけでなく広く「有機リン・カーバメート」としていることには留意されたい。たとえば野鳥については清宮ら(2007)[68]、水生生物は岩田(1981)[69]、菊地・若林(1997)[70]など。
ちなみに、同じく昆虫が病原体を媒介とするニレ立枯病においては病原菌媒介者であるキクイムシ類を標的としたDDT等の非選択性農薬の散布が行われたが、薬剤の生物濃縮による小鳥の中毒死など他生物に影響が出たので中止された。DDT禁止運動の発端になったのがレイチェル・カーソン著の有名な「沈黙の春」(原題:Silent Spring)であり、民衆を大きく動かした事例として知られる。
-
 フェニトロチオン(有機リン系殺虫剤)
フェニトロチオン(有機リン系殺虫剤) -
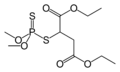 マラチオン(有機リン系)
マラチオン(有機リン系) -
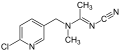 アセタミプリド(ネオニコチノイド系)
アセタミプリド(ネオニコチノイド系) -
 チアクロプリド(ネオニコチノイド系)
チアクロプリド(ネオニコチノイド系) -
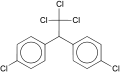 【参考】DDT(有機塩素系)
【参考】DDT(有機塩素系)





また、クロマツ、アカマツは、いずれも広葉樹の高木の生育が困難な厳しい環境条件下で局所的に安定した群落を維持することを除くと、広域で安定した森林を作らない。むしろ、植物遷移の上では、裸地に定着する先駆者樹木であり、林内が暗く富栄養になると次世代への更新ができない。そのため、松林を維持するには常に林床を貧栄養かつ明るい条件に維持する必要がある。かつて、松林の落葉落枝は広く燃料として利用され、そのために人里周辺では松林が維持されてきた。現在それが行われなくなったため、松林の減少するのは当然であり、マツ材線虫病は単にそれを加速したにすぎないという考えがある。 この考えに立てば、必要のなくなった松林では松くい虫防除は必ずしも行う必要がなく、むしろ殺虫剤の広域散布による弊害を考慮すべきである。用いられている殺虫剤は選択的にマツノマダラカミキリを殺すものではなく、森林の生物群集を構成する昆虫など地上生の節足動物を非選択的に殺すものであるために、森林の生態系を乱し多様性を損なう。農薬の人体への健康被害の研究と比べても、地域生態系への影響評価の研究はまだ十分とは言えず、この観点からの防除対策の評価が必要である。また、枯死した松を切り倒す作業による周囲の攪乱によって、自然植生の回復が遅れたり、被害が広がったと思われる例もある。
長野県松本市では、アカマツ林の松枯れ被害のうちマツ材線虫病による被害木の主原因とされるマツ材線虫を媒介伝播する主原因とされるマツノマダラカミキリの羽化後の成虫の駆除のみを目的としたニコチノイド系殺虫剤の無人ヘリコプターによる空中散布が2013年から行われているが、その効果は限定的で2019年現在散布地のアカマツ林の松枯れはさらに拡大もしくは枯損が拡がっている。このためドリフト等による健康被害や発達障害リスク税金補助金の無駄遣いを憂う複数の弁護士及び松本市民らによる反対署名活動や市街地での反対デモ行進など大規模な空中散布反対運動が展開され、2017年から松本市を被告とした訴訟がいくつか提起されている。この問題は国会でも取り上げられ、週刊誌やネットニュース等でも報道されている。[71][72][73][74]
批判への反論
[編集]農薬の空中散布に批判があるのは当然だが、それを理由に、あるいは別の理由で松くい虫防除や時にはマツ林そのものを不要とする意見が出されることもある。それらへの反論の一部を挙げる。
- すでに林野庁は方針を転換し、防除対象を特に守るべき林分(高度公益機能森林及び被害拡大防止森林)に限定している。現在防除対象となっているのは、理由があって守るべきと判断された林分である。従って農薬の無差別散布との批判は当たらない。
- 現在国や自治体によって行われている防除の手段は薬剤散布一辺倒ではなく、「特別伐倒駆除」及び「補完伐倒駆除」、「樹種転換」についても法律(森林病害虫等防除法)で定められている。
- 伝染病による急速なマツ林の崩壊は、正常な遷移の過程ではない。通常の陽樹から陰樹への遷移とは別の、元々生態系に存在しなかった要因による異常な現象であり、単なる「加速」とは言い切れない。
- 里山林としてのアカマツ林は、例えば京都の社寺における借景など景観の重要な要素ともなり、日本文化の中で無視できない位置を占める。アカマツ林は燃料などの供給源として成立したものだが、文化的観点からは単なる時代遅れの資源供給源として代替の効く存在ではない。その点においては生産手段としては必ずしも効率的ではない棚田と類似点がある。
- 海岸、特に砂丘のクロマツ林は必ずしも天然植生ではなく、その多くは古くは藩政期から(例:吹上浜)近くは昭和に至るまで(例:襟裳岬)防風防砂のために植栽されてきた人工林で、現在も飛砂防備保安林に指定されているものが多い。代替できる樹種がなく、その機能を維持するための防除が必要である。
歴史
[編集]マツ枯れによると考えられるマツの集団枯損の記録は、1905年に長崎市周辺で発生したものにまで遡る[75]。1914-15年頃には兵庫県赤穂市でも被害が発生し、以後九州山陽地方に被害が広がった。戦中戦後はマツ枯れ防除の余裕もなく、被害が拡大した。戦後はGHQの勧告のもと林野庁や各自治体が強力に伐倒駆除を進めた結果、一時被害は沈静化された。当時は家庭用燃料の3/4以上を薪が占めるなど燃料としての需要が旺盛だったため、マツ枯れ被害材も燃料用に飛ぶように売れたという。さらに里山アカマツ林では落枝落葉や球果(松かさ、松ぼっくり)も肥料や燃料として利用されたため、林内にはあまり有機物が堆積せず貧栄養に保たれ、先駆樹種であるアカマツに適した環境が維持されていた。
しかし1950年代の燃料革命や化学肥料の普及によって、肥料や燃料としてのマツ林の落枝落葉の需要が減少して林内に放置されたため、土壌が富栄養化してアカマツには不適当な環境になり始めた。マツタケの大幅減産もこの頃(1960年代前半)である。また、マツ材は強度が高いため屋根を支える梁などの建築用材としても生産・利用されてきたが、素材生産量は1957年の1551万立米をピークに減少に転じ、さらに1960年の木材輸入自由化後は価格競争にさらされ、1975年にはほぼ1/4にまで減少し、2004年にはわずか82万立米にまで落ち込んだ。このようなマツを巡る環境の下、防除の努力にもかかわらずマツ枯れ被害は1970年代から再び増加に転じ、1979年には被害材積が243万立米を記録している。以後毎年100万立米前後の被害が続いてきた。現在は保安林や景勝地などの保全と被害の北上を阻止することに重点をおいた防除が行われている。2015年現在、被害の北端は東北地方北部に達しており、日本海側が青森県南西部の深浦町付近、太平洋側はやや進行が遅く岩手県の中北部の八幡平市・滝沢市付近、三陸海岸沿いはさらに進行が遅く大船渡市付近となっている。
世界的な広がり
[編集]日本に近い東アジア諸国でもマツ枯れが問題になっている。中国では1982年に南京郊外で発生が確認され、バビショウ(タイワンアカマツ)や日本産クロマツに被害が発生している。台湾では1970年代からリュウキュウマツのマツ枯れが報告されている。香港や韓国[76]でも発生している。韓国では日本産アカマツやクロマツの被害が多かったが最近ではチョウセンゴヨウの枯死も確認され始めたという[77]。
ヨーロッパでは広く分布するヨーロッパアカマツ Pinus sylvestris やフランスカイガンショウ P. pinaster が感受性であることから、ヨーロッパ各国が北米からのマツ材の輸入を禁止する措置をとって貿易摩擦にまでなった。1999年にはポルトガルのリスボン近郊でマツ枯れの発生が確認され[78]、現在その封じ込めと根絶のための努力が行われている。
脚注
[編集]- ^ 日塔正俊, 小田久五, 加藤幸雄 (1967)、「522マツ類の穿孔虫に関する研究: 加害対象木の判定と季節的推移について(第78回日本林学会大会)」 『日本林学会誌』 1967年 49号 3号 p.126-126
- ^ 小田久五 (1967) 「松くい虫の加害対象木とその判定法について」 『森林防疫ニュース』16:263-266, NAID 10009947486
- ^ 笹川満廣・脇田棘三(1959)アカマツ穿孔ゾウムシ類の加害に関する一観察. 京都府立大學學術報告 農學11:71-74.
- ^ 徳重陽山, 清原友也(1969)マツ枯死木中に生息する線虫Bursaphelenchus sp. 『日本林學會誌』 51(7), 193-195.
- ^ a b 清原友也, 徳重陽山(1971)「マツ生立木に対する線虫Bursaphelenchus sp.の接種試験」 『日本林学会誌』 1971年 53巻 7号 p.210-218, doi:10.11519/jjfs1953.53.7_210
- ^ Mamiya, Y., and Kiyohara, T., (1972) Description of Bursaphelenchus lignicolus n. sp. (Nematoda: Aphelenchoidae) from pine wood and histopathlogy of nematode infected trees. Nematologica18(1) :120-124.
- ^ 真宮靖治, 清原友也, 徳重陽山(1971)「マツの材組織中にみられるBursaphelenchus属線虫の1新種, マツノザイセンチュウ(仮称)」 『日本応用動物昆虫学会大会講演要旨』 (15), 28
- ^ 山田利博, 伊藤進一郎 (1993)、「材線虫病抵抗性マツ,テーダマツおよびストローブマツ,のマツノザイセンチュウ感染に対する化学的防御反応」 『日本植物病理学会報』 1993年 59巻 6号 p.666-672, doi:10.3186/jjphytopath.59.666, 日本植物病理学会
- ^ 黒田慶子, 山田利博, 峯尾一彦, 田村弘忠、「マツ材線虫病の病徴進展におけるキャビテーションの影響」 『日本植物病理学会報』 1988年 54巻 5号 p.606-615, doi:10.3186/jjphytopath.54.606, 日本植物病理学会
- ^ 福田健二, 寳月岱造, 鈴木和夫、「病原性の異なるマツノザイセンチュウおよびニセマツノザイセンチユウ接種したクロマツ木部の解剖学的変化および通導阻害」『日本林學會誌』 1992年 74巻 4号 p.289-299, 日本林學會
- ^ 池田武文 (1994)「樹木に発生するキャビテーションのアコースティック・エミッション法による検出」 『日本林学会誌』 1994年 76巻 4号 p.364-366, doi:10.11519/jjfs1953.76.4_364, 日本森林学会
- ^ Steiner G., Burhrer E. M., (1934) Aphelenchoides xylophilus, n. sp., a nematode associated with blue-stain and other fungi in timber. Journal of Agriculture Research 48944951.
- ^ 堂園安生・清原友也 (1971) 菌糸培養法におけるマツノザイセンチュウの増殖温度. 日本森林学会九州支部研究論文 25: 160-161.
- ^ a b 清原友也 (1976)「マツノザイセンチュウの継代培養による病原性の低下」 『日本線虫学会誌』 1976年 6巻 p.56-59, doi:10.14855/jjn1972.6.56, 日本線虫学会
- ^ Kiyohara, T., and Bolla, R., I., (1990) "Pathogenic Variability Among Populations of the Pinewood Nematode, Bursaphelenchus Xylophilus.", Forest Science 36(4): 1061-1076.
- ^ 小島克己, 上條厚, 益守眞也, 佐々木惠彦 (1994)、「病原力の異なるマツノザイセンチュウの分泌するセルラーゼの性質」 『日本森林学会誌』 1994年 76巻 3号 p.258-262, doi:10.11519/jjfs1953.76.3_2587
- ^ a b c 森本桂, 岩崎厚、「マツノザイセンチュウ伝播者としてのマツノマダラカミキリの役割」 『日本林學會誌』 1972年 54巻 6号 p.177-183, doi:10.11519/jjfs1953.54.6_177, 日本森林学会
- ^ 真宮靖治(1980)、「マツノザイセンチュウの接種による当年生アカマツ苗の発病とその病態解剖」 『日本林学会誌』 1980年 62巻 5号 p.176-183, doi:10.11519/jjfs1953.62.5_176, 日本森林学会
- ^ a b 小林享夫, 佐々木克彦, 真宮靖治(1974)、「マツノザイセンチュウの生活環に関連する糸状菌 (I)」 『日本林學會誌』 1974年 56巻 4号 p.136-145, doi:10.11519/jjfs1953.56.4_136, 日本森林学会
- ^ 小林享夫・佐々木克彦・真宮靖治(1975)、「マツノザイセンチュウの生活環に関連する糸状菌 (II)」 『日本林學會誌』 1975年 57巻 6号 p.184-193, doi:10.11519/jjfs1953.57.6_184, 日本森林学会
- ^ 神崎菜摘, 二井一禎、「ニセマツノザイセンチュウのアカマツに対する病原性」 『日本応用動物昆虫学会大会講演要旨』 2003年 47巻 日本応用動物昆虫学会大会講演要旨, p.157, 日本応用動物昆虫学会
- ^ 石橋信義 編著. (2003) 線虫の生物学. 東京大学出版会. 東京.
- ^ a b 神崎菜摘, 竹本周平(2012)、「Bursaphelenchus属線虫の植物病原性と媒介者の生活史特性の関連」 『日本森林学会誌』 2012年 94巻 6号 p.299-306, doi:10.4005/jjfs.94.299, 日本森林学会
- ^ G. Skarmoutsos andH. Michalopoulos-Skarmoutsos (2000)Pathogenicity of Bursaphelenchus sexdentati, Bursaphelenchus leoni and Bursaphelenchus hellenicus on European pine seedlings. Forest pathology30(3) 149-156, doi:10.1046/j.1439-0329.2000.00198.x
- ^ 軸丸祥大(1996)、「カラフトヒゲナガカミキリの個体群動態とニセマツノザインチュウの伝播に関する研究 : 冷涼な地域におけるマツ材線虫病激害化の阻害過程」 『広島大学総合科学部紀要. IV, 理系編』 1996年 22巻 p.191-193, NCID AN10435936, 広島大学総合科学部
- ^ 越智鬼志夫(1969)、「マツ類を加害するカミキリムシ類の生態 (II) Monochamus属2種成虫の羽化と産卵習性などについて」 『日本林学会誌』 1969年 51巻 7号 p.188-192, doi:10.11519/jjfs1953.51.7_188, 日本森林学会
- ^ Togashi K., Taga Y., Iguchi, K., Aikawa T. (2008) Bursaphelenchus mucronatus (Nematoda : Aphelenchoididae) vectored by Monochamus urussovi (Coleoptera : Cerambycidae) in Hokkaido, Japan. Journal of Forest Research 13(2), 127-131, doi:10.1007/s10310-007-0057-1
- ^ 田中一二三, 玉泉幸一郎, 保坂武宣, (2008)、「根系癒合によるマツ材線虫病被害木の拡大」 『日本森林学会大会発表データベース』 第119回日本森林学会大会 セッションID: A09, doi:10.11519/jfsc.119.0.119.0, 日本森林学会
- ^ 黒田慶子, 伊藤進一郎(1992)、「クロマツに侵入後のマツノザイセンチュウの動きとその他の微生物相の変遷」 『日本林学会誌』 1992年 74巻 5号 p.383-389, doi:10.11519/jjfs1953.74.5_383, 日本森林学会
- ^ 作山健 (1985) マツつちくらげ病の発生生態と防除. 岩手県林業試験場報告18号
- ^ 佐藤邦彦, 横沢良憲, 庄司次男(1974)、「マツ類の群状枯死を起こす「つちくらげ」病に関する研究(「まつくいむしによるマツ類の枯損防止に関する研究」の一部)」『林業試験場研究報告』1974年 268号 p.13-48, 森林総合研究所
- ^ 中村克典, 秋葉満輝, 相川拓也, 小坂肇, 伊禮英毅, 喜友名朝次 (2010)、「沖縄県宮古島のリュウキュウマツ枯死木およびマツノマダラカミキリからのBursaphelenchus属線虫検出調査」 『日本森林学会誌』 2010年 92巻 1号 p.45-49, doi:10.4005/jjfs.92.45, 日本森林学会
- ^ 古野東洲(1982)、「外国産マツ属の虫害に関する研究 : 第7報 マツノザイセンチュウにより枯死したマツ属について」 『京都大学農学部演習林報告』 1982年 54巻 p.16-30, hdl:2433/191761, 京都大学農学部附属演習林
- ^ 二井一禎, 古野東洲(1979)、「マツノザイセンチュウに対するマツ属の抵抗性」 『京都大学農学部演習林報告』 1979年 51巻 p.23-36, hdl:/2433/191693, 京都大学農学部附属演習林
- ^ Akiba M., Nakamura K., (2005) Susceptibility of adult trees of the endangered species Pinus amamiana var. amamiana to pine wilt disease in the field. Journal of forest research 10(1): 3-7.
- ^ 金谷整一l, 中村克典, 秋葉満輝 ほか(2005)、「種子島木成国有林におけるマツ材線虫病で枯死したヤクタネゴヨウの伐倒駆除」 『保全生態学研究』 2005年 10巻 1号 p.77-84, NAID 110001888705, doi:10.18960/hozen.10.1_77, 日本生態学会
- ^ 岩手県林業技術センター(2005)ドイツトウヒに発生したマツ材線虫病. 林業技術情報No.21
- ^ 藤田恵美, 中田誠 (2001)、「海岸砂丘地のクロマツ林における広葉樹の混交による立地環境の変化 新潟県下越地方の事例」 『日本森林学会誌』 2001年 83巻 2号 p.84-92, doi:10.11519/jjfs1953.83.2_84
- ^ 中村充博, 鈴木祥悟(2007)、「アカゲラ誘致のための人工巣丸太の架設実験」 『総合政策』 2007年 9巻 1号 p.49-58, 森林総合研究所東北支所
- ^ 岡本安順(1999)、「マツノマダラカミキリの天敵昆虫サビマダラオオホソカタムシの寄生状況と生態調査」 『森林応用研究』 1999年 8巻 p.229-232, doi:10.20660/applfor.8.0_229, 応用森林学会
- ^ 喜友名朝次(2009)、「松くい虫天敵昆虫防除技術開発研究」『沖縄県森林資源研究センター研究報告』 2011年 52号 p.12-15, NAID 120005321945, 沖縄県森林資源研究センター
- ^ 牧本卓史(2011)、「サビマダラオオホソカタムシの松くい虫防除への適用」 『岡山県農林水産総合センター研究報告』 2011年 27号 p.73-82, NAID 120005330890, 岡山県農林水産総合センター森林研究所
- ^ 上田明良, 藤田和幸, 浦野忠久, 山田倫章(1999)、「オオコクヌスト成虫放虫によるマツノマダラカミキリ捕食実験」 『森林応用研究』 1999年 8巻 p.169-172, doi:10.20660/applfor.8.0_169, 応用森林学会
- ^ 岡本安順(1997)、「スギカミキリ・マツノマダラカミキリに対するクロアリガタバチの寄生実験」 『森林応用研究』 1997年 6巻 p.131-134, doi:10.20660/applfor.6.0_131, 応用森林学会
- ^ H. Kosaka, and N. Ogura, (1993) "Contortylenchus genitalicola n.sp.(Tylenchida:Allantonematidae)from the Japanese Pine Sawyer, Monochamus alternatus(Coleoptera:Cerambycidae)". 『日本応用動物昆虫学会誌』 1993年 28巻 4号 P.423-432, 日本応用動物昆虫学会
- ^ Bedding R. A. (1968) Deladenus wilsoni n. sp. and D. siricidicola n. sp. (Neotylenchidae), entomophagous-mycetophagous nematodes parasitic in siricid woodwasps. Nematologica 14: 515-525.
- ^ Bedding, R.A., and Akhurst R. J., (1971)Use of the nematode Deladenus siridicola in the biological control of Sirex noctilio in Australia. Australian Journal of Entomology 13: 129-135.
- ^ Sayama K., Kosaka H., and Makino, S., (2007) The first record of infection and sterilization by the nematode Sphaerularia in hornets (Hymenoptera, Vespdiae, Vespa). Insectes Sociaux54: 53-55.
- ^ 相川拓也(2012)、「マツノマダラカミキリに残る共生細菌感染の痕跡」『日本森林学会誌』 2012年 94巻 6号 p.292-298, doi:10.4005/jjfs.94.292, 日本森林学会
- ^ 上田明良(1998)、「マダラコールを用いた誘因トラップの設置高別捕獲調査」 『森林応用研究』 1998年 7巻 p.109-112, doi:10.20660/applfor.7.0_109, 応用森林学会
- ^ 高梨琢磨.・小島渉(2012)森林昆虫における振動情報の機能解明と害虫防除への応用. 木材保存38(4):157-162.
- ^ Muramoto M. ,(1999) Ending of pine wilt disease in Okinoerabu island, Kagoshima prefecture. In Futai, K., Togashi, K. & Ikeda, T. eds. Sustainability of Pine Forests in Relation to Pine Wilt and Decline. Shokado shoten, Kyoto.
- ^ Nakamura K., Yoshida N., (2004) Successful control of pine wilt disease in Fukiage-hama seacoast pine forest in southwestern Japan. In Mota M., Viera P., eds. The pine wood nematode Bursaphelenchus xylophilus. proceeding international workshop, University of Evora, Portugal August 2001.
- ^ 松浦邦昭・細野隆次(1987)マツノザイセンチュウのアセチルコリンエステラーゼの特性と防除剤によるその活性阻害. 日本線虫学会誌17: 17-22.
- ^ 小倉信夫(2004)培養条件下でのマツノザイセンチュウの増殖に対する数種殺虫剤の抑制効果. 日本線虫学会誌34(2): 99-102
- ^ 竹下努 (1992) マツ材線虫病の注入傷害防止に関する研究. 鳥取県林業試験場研究報告第34号.
- ^ 中川茂子 (2001) マツ枯れ防止樹幹注入剤施用により発生する異常. 樹木医学研究5(1): 13-20.
- ^ 真宮靖治・平塚美幸・村田政穂(2005)木材腐朽菌のマツノザイセンチュウに対する捕食効果. 日本線虫学会誌 35(1): 21-30.
- ^ 清原友也 (1990) マツ材線虫病における誘導抵抗性. 日本農芸化学会誌64(7): 1251-1253.
- ^ B. S. Griffiths, S. Caul, J. Thompson, A. N. E. Birch, C. Scrimgeour, M.N. Anderson, J. Corter, A. Messean, C. Sausse, B. Lacroix, P.H. Krogh, (2005) A comparison of soil microbial community structure, protozoa and nematodes in field plots of conventional and genetically modified maize expreessing the Bacillus thuringiensis Cry1Ab toxin. Plant and soil 275: 135-146.
- ^ “森林病害虫等防除法(昭和二十五年法律第五十三号)第3条第5項:駆除命令、移動を制限”. e-Gov法令検索. 総務省行政管理局 (2016年5月20日). 2020年1月15日閲覧。 “2017年4月1日施行分”
- ^ 千葉春美・石井幸男・富岡甲子次・飯塚三男.(1963)赤沼におけるテーダマツの成長. 林業試験場報告161号.
- ^ 中井勇・光枝和夫・大畠誠一 (1995) マツ属の種間交雑からみた種の類縁関係とF1雑種の生育状況について. 京都大学農学部演習林報告67: 1-18.
- ^ 後藤晋・宮原文彦・井出雄二(2002) クロマツ種苗のマツ材線虫病抵抗性に対する花粉親の寄与 日本林学会誌
- ^ 石川広隆(1968) マツ・カラマツ類を中心としたさし木困難樹種の不定根の形成に関する基礎研究 1968 林試研報214号
- ^ 後藤晋 (1999) クロマツの挿し木増殖における発根条件の検討 日林九支研論52号
- ^ 松永孝治・大平峰子・倉本哲嗣(2009) さし穂サイズと採穂台木の形態的要因がクロマツさし木苗の生産効率に与える影響.日本森林学会誌91(5): 335-343.
- ^ 清宮幸男・古川岳大・高橋真紀・田村貴・村上隆宏・関慶久・佐々木幸治 (2007)野鳥のコリンエステラーゼ阻害剤中毒. 日本獣医師会雑誌
- ^ 岩田治郎(1981)農薬の沿岸生態系への影響(シンポジウム:沿岸海域における人工有機化合物汚染). 沿岸海洋研究ノート19(1):47-55.
- ^ 菊地幹夫・若林明子(1997)オオミジンコによる河川水中の化学物質の有害性モニタリング. 日本水産学会誌. 63(4):627-633.
- ^ “松本の松枯れを考える住民の会”. www.facebook.com. 2019年5月26日閲覧。
- ^ “発達障害リスク「農薬」に対策遅れる日本 禁止どころか緩和も 〈週刊朝日〉”. AERA dot. (アエラドット) (2019年3月6日). 2019年5月26日閲覧。
- ^ “松本の松枯れを考える住民の会”. www.facebook.com. 2019年5月26日閲覧。
- ^ “ネオニコチノイド系農薬等に関する質問主意書”. www.shugiin.go.jp. 2019年5月26日閲覧。
- ^ 矢野宗幹(1913)長崎県下松樹枯死原因調査. 山林公報4
- ^ S. C. Shin, (2008) Pine wilt disease in Korea. Pine Wilt Disease 26-32.
- ^ H. Han, Y.-J. Chung, and S.-C. Shin,(2008) First report of pine wilt disease on Pinus koraiensis in Korea. Plant Disease 92(8): 1251-1251.
- ^ M. M. Mota; H. Braasch; M. A. Bravo; A. C. Penas; W. Burgermeister; K. Metge; E. Sousa,(1999)First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1(7): 727-734.
参考文献
[編集]- マツ枯れは森の感染症 森林微生物相互関係論ノート 二井一禎 (2003) 文一総合出版 ISBN 4-8299-2183-8
- 森林微生物生態学 二井一禎・肘井直樹 編著 (2000) 4.5 微生物と線虫を利用する昆虫の繁殖戦略―マツノマダラカミキリによるマツノザイセンチュウの伝播― 前原紀敏 朝倉書店 ISBN 4-254-47031-2
- きのこの自然誌(新装版) 小川真 (1983) 築地書館 ISBN 978-4-8067-2198-7
- 林業普及シリーズ第36集 家庭燃料の話 奥田富子・内田憲 (1953) 林野庁指導部研究普及課(非売品)
- 農林省統計表(農林水産省統計表)第31次〜第80次 農林省(農林水産省) 農林統計協会
関連項目
[編集]樹木の世界三大(四大)病害
[編集]- 五葉マツ類発疹さび病 - アジア産の病原菌がアメリカのマツに壊滅的被害を与えている病気。
- クリ胴枯病 - アジア産の病原菌が欧米、特にアメリカのクリに壊滅的被害を与えている病気。
- ニレ立枯病(オランダニレ病) - アジア産の病原菌が欧米のニレに壊滅的被害を与えている病気。道管の閉塞による通水障害と病原体を昆虫が媒介するという点は似ている。
- マツ材線虫病 - 本項で解説。
マツ・針葉樹の重大な病気
[編集]- 五葉マツ類発疹さび病 - アメリカで五葉マツ類を植える時最も障害になる病気。上述。
- マツ材線虫病 - 日本でマツを植える時最も障害になる病気。本項で解説
- スギ赤枯病 - スギ苗木を枯死に至らせる重要病害で成木にも感染しスギ溝腐病を引き起こし木材の経済的価値を落とす。病原菌はアメリカ原産とされる。
- 根腐れ - 原因菌は複数知られるが特にナラタケ類、マツノネクチタケ類(特に欧米産)、エキビョウキン類などによるものが成木でも枯死被害が大きく有名
- 心材部の腐朽 - 外見は正常だが木材の強度が下がり倒れやすくなることで間接的な死因になる。原因菌は複数知られる。
穿孔性昆虫やその共生生物による樹木の主な病気
[編集]- ニレ立枯病(オランダニレ病) - キクイムシが病原菌を媒介
- ナラ枯れ - ナガキクイムシが病原菌を媒介
- 北米マツ属樹木に対するに対するDendroctonus属キクイムシの被害
- モミ属・カラマツ属樹木に対するヤツバキクイムシ類の被害
- ヤシの赤輪病 - ゾウムシが病原線虫を媒介
- オセアニア・北米のマツ属樹木に対するノクチリオキバチによる被害
- アメリカのトネリコ属樹木に対するアオナガタマムシによる被害
生物
[編集]マツと環境
[編集]外部リンク
[編集]関連生物・病気
[編集]- 森林生物00154 マツノザイセンチュウ 森林総合研究所資料
- 森林生物00155 マツノマダラカミキリ
- 森林生物00253 カラフトヒゲナガカミキリ
- 森林生物00264 オオコクヌスト カミキリの天敵として期待される昆虫
- 森林生物00284 サビマダラオオホソカタムシ 同上
- マツ枯れ…マツ材線虫病 神戸大学森林資源学研究室
- Pine wilt disease ― American Phytopathological Society
- How to Identify and Manage Pine Wilt Disease and Treat Wood Products Infested by the Pinewood Nematodes
- red ring nematode - Bursaphelenchus cocophilus Cobb 近縁線虫が引き起こすヤシの赤輪病についてフロリダ大学
技術
[編集]- アコースティックエミッションによるトマトの水ストレスの検出 農研機構HPによるAE法の応用事例紹介
薬剤
[編集]- 殺虫剤樹木散布編 東京都環境局 化学物質の子どもガイドライン
法律
[編集]- 森林病害虫等防除法(昭和二十五年法律第五十三号).e-Gov法令検索. 総務省行政管理局
| 典拠管理データベース: 国立図書館 |
|---|