
ウサロミケス
| ウサロミケス | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Utharomyces epallocaulus
| ||||||||||||||||||
| 分類 | ||||||||||||||||||
| ||||||||||||||||||
| 学名 | ||||||||||||||||||
| Utharomyces Boedijn, 1958 | ||||||||||||||||||
| 種 | ||||||||||||||||||
|
本文参照 |
ウサロミケス Utharomyces はケカビ目のカビの1つ。ミズタマカビに似て、動物の糞上に生育し、長く伸びる胞子嚢の先端には硬化した胞子嚢をつけ、その直下が膨らんで頂嚢となる。ただし胞子嚢を打ち出すことはせず、器物に触れた際に頂嚢が壊れることで胞子嚢をそこに貼り付ける。
概説
[編集]ウサロミケスはミズタマカビとよく似た特徴を持つカビで、基質菌糸に作られる栄養嚢から伸び出した胞子嚢柄の先端に胞子嚢を形成し、そのすぐ下の部分が膨大して頂嚢を作ること、胞子嚢壁が壊れず、胞子嚢が胞子嚢胞子をその中に抱えたままで外れることが共通している。ただしミズタマカビでは胞子嚢が強く打ち出されるが、このカビはそのようなことがない。またミズタマカビは動物の糞に出て、糞に特有の成分がなければ発育しないのに対して、この菌はやはり糞に出るものの特殊な成分を要求することはない。このような点から、本菌はミズタマカビ同様に糞生菌としての特殊化が強く見られるものであるが、ミズタマカビほどそれが強くないもの、と判断されてきた。熱帯から亜熱帯に見られるもので、その限りでは世界に広く分布している。
特徴
[編集]
タイプ種である U. epallocaulus に基づいて記す[1]。
栄養体
[編集]腐生菌であり、通常の培地で培養が可能である。その成長は速く、麦芽寒天培地の上に20℃で培養した場合、径8.5cmのペトリ皿を10日で完全に覆い尽くす。培地より上に出てくるのは胞子嚢柄のみで、気中菌糸は全く出さない。
無性生殖
[編集]無性生殖は胞子嚢胞子による。基質中の菌糸にはまずその先端に、あるいは介在的に生じた栄養嚢(Trophosyst)が形成され、これは寒天中に作られるもので亜球形、幅広い楕円形、卵形などの形をしている。胞子嚢柄はこの栄養嚢から伸び出す。胞子嚢柄は当初は上向きに直立して伸び、次第にやや斜めに伸び、その後には何かにもたれかかって伸びる。その伸長は早く、長さは2cm、あるいはそれ以上にまで伸び、その径は8-20μmほど。胞子嚢柄は正の光屈性を示す。胞子嚢柄は分枝をせず、その先端にたった一つだけの胞子嚢を付ける。胞子嚢はその形成の初めには球形で白いが、成熟時には何となく半球形になり、黒くてその表面は光を反射するようになる。表面には細かな棘状突起が並び、大きさは75-180×65-150μm。胞子嚢柄の伸長が終わり、胞子嚢が成熟すると、幅広い楕円形の、薄い壁から成る頂嚢が胞子嚢柄の壁の拡張と引き延ばしの結果として生じ、その大きさは径80-200μmほどとなり、位置は胞子嚢からわずかだけ距離を置いたすぐ下である。それから胞子嚢直下の柄、胞子嚢柄に形成された頂嚢と胞子嚢の間の胞子嚢柄の部分、長さにして35-130μmの部分、先細りになっていて基部の太さが18-36μm、胞子嚢直下の太さが12-18μmの部分の細胞壁が厚くなり、厚さ5μmにまでなるという形で、この部分の形成が頂嚢形成の後に行われる。柱軸は上部が球形に膨らみ、その下で強く絞られており、さらにその下が半球形に膨らんでいて、上の段の膨らみが24-60×16-40μm、下の段の膨らみが16-34×12-20μm。薄膜で出来ており、早くに崩れる。胞子嚢胞子は平らな円柱状をしており、径6.5-7μm、厚みが4.5-5μm。両極の面の中央付近および側面を巡る帯状部分は明らかに小突起が密生しており、やや無色であるがまとまると灰色を帯びる。
-
 完成した胞子嚢の部分
完成した胞子嚢の部分 -
 頂嚢の上半と胞子嚢が外れた状態
頂嚢の上半と胞子嚢が外れた状態 -
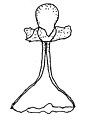 胞子嚢壁を壊して柱軸を見せた状態
胞子嚢壁を壊して柱軸を見せた状態 -
 胞子嚢胞子
胞子嚢胞子




有性生殖
[編集]有性生殖は配偶子嚢接合によるものと思われるが、観察されてはいない。自家不和合性であると推察される。1980年時点では接合胞子嚢を得るための試みは何度か行われているもののすべて失敗している[2]。
-
 胞子嚢の形成過程
胞子嚢の形成過程
気質菌糸の栄養嚢から伸張が始まる -
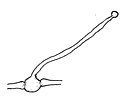 胞子嚢柄が伸び、先端に胞子嚢が作られ始める
胞子嚢柄が伸び、先端に胞子嚢が作られ始める -
 胞子嚢が膨らむ
胞子嚢が膨らむ -
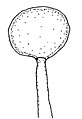 胞子嚢がさらに膨らむ
胞子嚢がさらに膨らむ -
 胞子嚢が色づく
胞子嚢が色づく -
 胞子嚢下に頂嚢が膨らみ始める
胞子嚢下に頂嚢が膨らみ始める






生育環境
[編集]自然界では糞に固有の糞生菌と考えられ、分離源となっているのは動物の糞がほとんどではあるが、土壌や腐植質から発見された例もある[3]。
胞子散布の様式
[編集]成熟した胞子嚢の表面は細かな棘状突起のために疎水性を持ち、他方でそれより下の胞子嚢を支える細い柄や頂嚢の部分はそうなっていないので親水的である[4]。そのために胞子嚢が湿った面に押し当てられた場合、その疎水性のために反発力を生じ、胞子嚢柄や頂嚢がそこに触れやすくなるように向きを変えさせ、おそらくは頂嚢を破裂させるに足る力を加えさせる。それによって頂嚢の内容物が放出され、それが乾燥することでその部分より先の部分、つまり頂嚢の上半分と細い柄と胞子嚢がその部分に固着されることとなる。胞子嚢胞子は胞子嚢が機械的に破壊された場合にのみ放出される。
分布
[編集]発見数は多くないものの、その地域はアフリカのガーナ、インドネシアのジャワ、インドのラジャスタン、北アメリカのフロリダと熱帯域としては世界全体にわたっている[3]。なお、この種は日本でも発見例があり、東京新宿のネズミ糞から報告されている[5]。
分類
[編集]本種はその形態がミズタマカビ Pilobolus によく似ており、ごく近縁なものと考えられた[6]。そのために本属を記載した Boedijn はミズタマカビ科 Pilobolaceae に属するものと見なし、他の研究者もそれを疑うことはなかった[3]。ここには同時にピライラ Pilaira が含められており、この3属は非常によくまとまった自然群をなすと考えられた。より詳しく見ると、糞生菌として高度の適応を遂げたミズタマカビと、その方向への進化の初期段階を示すピライラに対して、本属はその両者の中間の段階を示すものとされた。つまり、まずこの三者は胞子嚢壁が分解せず、胞子嚢胞子をその中に含んでまとめたままに胞子を散布する点で共通し、これは糞上から周囲の植物体上に胞子を散布し、草食動物がそれを食べて糞に出すことで生育地を確保する、という糞生菌としての適応である。その中でミズタマカビは胞子嚢柄を栄養嚢上に出し、伸ばした胞子嚢柄の先端に頂嚢を生じ、これが破裂することで胞子嚢を射出する。またこの属のものは栄養要求が糞に特化し、糞特有の成分がなければ生育できない。対してピライラは栄養嚢も頂嚢も形成せず、胞子嚢柄はより長く伸長し、直接に胞子嚢を押し当てるようにして散布する。また生育に糞の成分を必ずしも必要としない。本属は栄養嚢と頂嚢を形成するが、胞子嚢を射出することはなく、胞子嚢柄は長く伸びる。栄養的には糞の成分を必要とはしない。これは確かに両者の中間と見える。
しかし、このような形態による分類が類縁関係を反映しないことが分子系統の研究で示され、ケカビ目の分類体系は全面的に変更され、ほとんどの科がばらばらになる惨状を呈した。その中で本群に関しては従来の見解が大きく維持された希なケースとなっている[7]。それによると、本属はやはりミズタマカビ属とごく近縁であり、この2属で1つのクレードを作ること、そしてピライラだけは大きく離れた系統のものであることが示された。この研究ではミズタマカビ属は複数の種が用いられているが、それらは1つのまとまった群となり、その群に対して本属が姉妹群を形成している、という理想的な結果が得られており、これら2属をまとめてミズタマカビ科とすることが提起されている。なお、この科ともっとも近い位置にあるとされているのはバクセラ属 Backusella のみを含むバクセラ科 Backusellaceae となっている。
下位分類
[編集]タイプ種の記載の後、新たな種として U. indica が1974年に記載された[8]。タイプ種より小型で、また柱軸や胞子嚢胞子の形が異なるものとしていたが、後の研究者はこれをタイプ種の範囲の中と判断している。現時点で本属はタイプ種のみの単形属とされる。
出典
[編集]- ^ 以下、記載は主としてKirk & Benny(1980)p.124-125
- ^ Kirk & Benny(1980)p.130
- ^ a b c Kirk & Benny(1980)p.126
- ^ 以下、Kirk & Benny(1980)p.127-130
- ^ Mikawa(1979)
- ^ 以下、Kirk & Benny(1980)p.123
- ^ 以下、Hoffmann et al.(2013),p.71
- ^ Sarbhoy et al.(1974)
参考文献
[編集]- P. M. Kirk & G. L. Benny, 1980. The genus Utharomyces Boedijn (Polobolaceae: Zygomycetes). Trans. Brit. mycol. Soc. 75(1) :p.123-131.
- A. K. Sarbhoy et al. 1974. An Interesting Member of the Pilobolaceae. Trans. Brit. mycol. Soc. 62(2) :p.425-427.
- K. Hoffmann et al. 2013. The family structure of the Mucorales: a synoptic revision based on comprehensive multigene-genealogies. Persoonia 30:p.57-76.
- Takashi Mikawa, 1979. A taxonomic study on Japanese sporangiferous Mucorales (5). Journ. Jap. Bot. Vol.54 No.7 :p.193-203.