
マツ科
| マツ科 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 ヨーロッパアカマツ
| |||||||||||||||
| 分類 | |||||||||||||||
| |||||||||||||||
| 学名 | |||||||||||||||
| Pinaceae Lindley | |||||||||||||||
| 和名 | |||||||||||||||
| マツ科 | |||||||||||||||
| 属 | |||||||||||||||
|
マツ科(マツか、学名:Pinaceae)は、裸子植物であり、球果植物(マツ門)に属する科である。北半球を代表する針葉樹のグループであり、針葉樹では現在最も繁栄しているとみられている一群である。ちなみに南半球ではマツ科ではなく、マキ科(Podocarpaceae)とナンヨウスギ科(Araucariaceae)の分化が著しく、ヒノキ科(Cupressaceae)は両半球の温帯地域に分布する。
形態
[編集]樹形はクリスマスツリーのような円錐形になるものが多い。枝は同じ高さから四方八方に伸ばすものが多い(輪生という)。枝には二種類あり、旺盛に伸長し我々が一般に「枝」と呼ぶものを「長枝」、葉の付け根にあるごく短いものを「短枝」と呼ぶ。長枝はすべての種が持つが、短枝は一部の属しか持たない。
-
 マツ属の樹形の例
マツ属の樹形の例 -
 カラマツ属の樹形の例
カラマツ属の樹形の例 -
 ヒマラヤスギ属の樹形の例
ヒマラヤスギ属の樹形の例 -
 トウヒ属の枝の付き方。同じ高さから枝を伸ばす
トウヒ属の枝の付き方。同じ高さから枝を伸ばす


,_Djebel_Chelia,_Algeria_1.jpg)

葉は針状であり生える場所は、長枝からのみ生えるもの(モミ属など)、長枝と短枝の両方から生えるもの(カラマツ属、ヒマラヤスギ属など)、短枝からしか生えないもの(マツ属)に分けられる。長枝から生える葉は一本ずつであるが、短枝から生える葉は数本から数十本が束になって生える。落葉の時にも特徴があり、マツ科では葉が落ちるときは葉だけが落ちる構造になっている。これに対し、マツ科と並んで代表的な針葉樹のグループであるヒノキ科では枝の一部分も一緒に落ちる。
-
 長枝からのみ葉を伸ばすトウヒ属
長枝からのみ葉を伸ばすトウヒ属 -
 短枝の先端から葉が束になって生えるカラマツ属
短枝の先端から葉が束になって生えるカラマツ属 -
 長枝にも短枝にも葉を付けるヒマラヤスギ属
長枝にも短枝にも葉を付けるヒマラヤスギ属 -
 参考:ヒノキ科のメタセコイアの落ち葉
参考:ヒノキ科のメタセコイアの落ち葉




花は雌雄同株(雄花と雌花が同じ株につくこと)、雄花は単生もしくは多数が束生する。花粉は風媒され、一般に2つの気嚢を持つが、カラマツ属、ヒマラヤスギ属、トガサワラ属、ツガ属では気嚢は1つである。雌花および雌花が成長した球果は鱗片状の構造が多数集まったものである。マツ属のみ原則として受粉した翌年に熟すが、ほかの種類は受粉した年に成熟する。また、マツ属のみ球果の鱗片が肥大して成長し、突起状の構造物(英:umbo)というものができる。熟した球果はそのまま落下するもの(マツ属、カラマツ属、トウヒ属、ツガ属、ユサン属など)と樹上で鱗片が分解するもの(モミ属、ヒマラヤスギ属、イヌカラマツ属など)がある。種子は一般に翼をもち、風で飛ばされやすい形となっているが、マツ属の一部には翼を持たないものがある。種子の翼は片側から伸ばす。ヒノキ科では種子の両側に翼がつく
-
 多数の雄花が集まるマツ属
多数の雄花が集まるマツ属 -
 トウヒ属の雄花
トウヒ属の雄花 -
 イヌカラマツ属の雄花
イヌカラマツ属の雄花 -
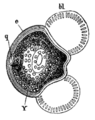 マツ属の花粉は気嚢を2つ持つ
マツ属の花粉は気嚢を2つ持つ -
 トウヒ属の種子
トウヒ属の種子 -
 参考:ヒノキ科ローソンヒノキの種子
参考:ヒノキ科ローソンヒノキの種子






生態
[編集]常緑樹が大半であるが、カラマツ属など落葉樹のグループもある。山火事が頻発するような地域に生えるマツ属の一部の種類には晩生球果(英:serotinous cone)と呼ばれる仕組みを持つものがあり、成熟しても球果の鱗片は開かないが、山火事の強熱を浴びたときに鱗片が開いて種子を飛ばす。山火事後の森林は上層木が焼かれて明るく、土壌中の病原菌や競合種の種子も壊滅しているので、光や栄養をめぐる生存競争に優位に立っていると考えられている。山火事の利用はマツ科ではマツ属の一部の種にしか知られていないが、南半球を中心にヒノキ科、フトモモ科、ヤマモガシ科など複数の科の一部の種で似たような生存戦略を持つものが知られる。
マツ科では苗木のうちの病原菌を避ける手段として山火事ではなく、倒木を利用する者もいる。落ち葉が積もった地上よりも倒木の上で発芽し成長したほうが、病原菌に侵される危険が少なく生存率が高い。
-
 晩生球果を持つマツの一種 Pinus banksiana
晩生球果を持つマツの一種 Pinus banksiana -
 山火事後に一斉に生えてきたマツ属Pinus contorta
山火事後に一斉に生えてきたマツ属Pinus contorta


マツ科の樹木はベニタケ科、フウセンタケ科、イグチ科、キシメジ科などの多くの菌類と菌根を形成する共生関係にある。樹木にとっては菌根を形成することで、土壌中の栄養分の吸収促進や菌類が作り出す抗生物質等による病原微生物の駆除等の利点があり、菌類にとっては樹木から光合成産物の一部を分けてもらうことができる。土壌中には菌根から菌糸を介し同種他個体や他種植物に繋がる広大なネットワークが存在すると考えられている[1][2][3][4][5][6]。共生する菌類の子実体は人間がキノコとして認識できる大きさに育つものが多く、中には食用にできるものもある。
-
 トウヒ属樹木の根(茶色の部分)と根を取り囲む菌類(白色の部分)
トウヒ属樹木の根(茶色の部分)と根を取り囲む菌類(白色の部分) -
 菌根が形成されたトガサワラ属樹木の根
菌根が形成されたトガサワラ属樹木の根 -
 菌根によるネットワークの模式図
菌根によるネットワークの模式図 -
 マツ属樹木下に発生したハツタケ(ベニタケ科)
マツ属樹木下に発生したハツタケ(ベニタケ科) -
 カラマツ属樹木と共生を結ぶハナイグチ(イグチ科)
カラマツ属樹木と共生を結ぶハナイグチ(イグチ科) -
 ショウゲンジ(フウセンタケ科)
ショウゲンジ(フウセンタケ科)





_Fr_473941.jpg)
人間との関わり
[編集]象徴
[編集]落葉広葉樹林が主体となるような地域(温帯など)においては、冬でも青々とした葉を付けるマツ科の常緑樹は生命力の象徴とされることがある。たとえば中国文化圏における歳寒三友(日本では主に松竹梅と呼ばれる)ではマツ属の樹木が崇められている。欧米ではモミ属やトウヒ属が魔除けに用いられることもあるという。クリスマスのシンボルであるクリスマスツリーになるのも円錐形の樹形を持つマツ科の常緑樹であることが多い。モミ属やトウヒ属が用いられることが多いが、これらは分布域が比較的狭いためより広い分布と温度適性を持つマツ属を使ったツリーもあるという。
-
 冬枯れの中でも目立つモミ属。
冬枯れの中でも目立つモミ属。 -
 クリスマスツリーとして販売されるモミ属
クリスマスツリーとして販売されるモミ属


木材
[編集]ヒノキ科などの他の針葉樹のグループと比べてより広い分布域を持つマツ科樹木は世界各地で木材としてよく利用される。マツ科樹木の天然分布は北半球のみであるが、南半球の地域でも大規模に植栽されていることがしばしばある。たとえばニュージーランドではアメリカ原産のラジアータマツ(Pinus radiata)に全面的に頼る林業を行っていることでよく知られている。日本の本州以南ではヒノキ科針葉樹の生育に向くことからマツ属のアカマツ(Pinus densiflora)を除いて木材目的での植栽は行われることが少ない。アカマツにしてもヒノキ科と比べたときに樹形の不安定さによる歩留まりの悪さ、マツ材線虫病(松くい虫、pine wilt)という致命的な伝染病の国内での流行に伴い新規植栽面積は激減している。国産アカマツ材の主要産地である岩手県三陸地方では超高密度植栽によって、通直で製材した時に歩留まりの良い樹形を作るなど造林技術を持つ地域もある。寒冷地である北海道は道南の一部を除いてヒノキ科針葉樹やマツ科マツ属ではなく、針葉樹の林業と言えばマツ科モミ属のトドマツかトウヒ属のアカエゾマツ、エゾマツで行われる。
木材のSPF材はトウヒ(英:Spruce)、マツ(英:Pine)、モミ(英:Fir)の頭文字を並べたもので、いずれも分類学上はマツ科に含まれる。ヒノキ科樹木が分布しないヨーロッパやカナダ、ヒノキ科樹木が分布するがマツ科樹木の利用が盛んなアメリカなどからの輸入材でホームセンターなどで手ごろな値段で手に入れることができる。
マツ属やカラマツ属の木材は心材と辺材の区別が明瞭であるが、トウヒ属やモミ属では不明瞭で区別できない。後者のような木材を淡色心材や熟材と呼ぶ。
-
 木材として利用されるマツ属
木材として利用されるマツ属 -
 心材と辺材の区別が明瞭なマツ属木材
心材と辺材の区別が明瞭なマツ属木材 -
 心材と辺材の区別が不明瞭なモミ属木材
心材と辺材の区別が不明瞭なモミ属木材



防災・並木
[編集]マツ属樹木を中心に海岸付近や荒れ地の緑化に使われることがある。特に強風による飛砂被害を軽減するため、また数mの盛土工と合わせて高潮・津波被害を軽減するために造成される。日本では山形県庄内地方で江戸時代から植栽が行われた記録が残るほか、全国各地に「松原」などと呼ばれ点在する。現在の日本においては林野庁もしくは国土交通省、もしくはこれらの省庁が管轄する都道府県の部署が治山事業や砂防事業によって松原の造成や管理を担当している。保安林や砂防指定地による伐採及び開発規制がかかっている場所が多い。トウヒ属は防雪林にもちいられることもある。
マツ属やヒマラヤスギ属はしばしば街路樹としても用いられ並木道を作るが、比較的古いものが多く新規では稀である。
-
 庄内砂丘(山形県、飛砂防備保安林)
庄内砂丘(山形県、飛砂防備保安林) -
 気比松原(福井県、潮害防備保安林)
気比松原(福井県、潮害防備保安林) -
 奇跡の一本松(岩手県、潮害防備保安林)
奇跡の一本松(岩手県、潮害防備保安林) -
 湘南海岸のマツ林(神奈川県、砂防指定地)
湘南海岸のマツ林(神奈川県、砂防指定地) -
 アッピア街道の松並木(イタリア)
アッピア街道の松並木(イタリア) -
 御油の松並木(愛知県)
御油の松並木(愛知県)






樹脂
[編集]マツ科樹木は細胞中に大きな樹脂道を持ち、樹脂の量が多く各地で利用されている。特にマツ属とモミ属でよく知られている。
食料
[編集]マツ属の一部の種の種子は松の実(英:pine nut)などと呼ばれ各地で食用にされる。樹木の直接の産物ではないが前述のように菌根を形成することから、マツ科樹木を中心とする森林では菌根性のキノコがよく発生し、その周辺地域では食毒問わずキノコに方言名を付けたり独自の食文化が発達していることもある。
-
 食用のために殻付きで乾燥させているマツ属の種子
食用のために殻付きで乾燥させているマツ属の種子 -
 殻を割って取り出したマツ属の種子
殻を割って取り出したマツ属の種子 -

-
 市場に並ぶアカハツタケ近縁種(スペイン)
市場に並ぶアカハツタケ近縁種(スペイン)




精油
[編集]葉から精油が採れる[7]。精油に含まれる香りの主成分は酢酸ボルニルであり、いわゆる「森の香り」がする[7]。これを利用して、「ウッディな香り」がする石鹸・入浴剤・ルームフレグランスなどが製造され、男性用化粧品にも多く使われる[7]。
花粉症の原因として
[編集]症例は少ないとされるが、花粉症の原因植物となることがある[8]。
-
 マツの葉を編んで作られた籠
マツの葉を編んで作られた籠 -
 マツ属の種子を混ぜたソース
マツ属の種子を混ぜたソース -
 ヒマラヤスギ属の精油
ヒマラヤスギ属の精油



分類
[編集]形態学的な分類で4亜科に分かれる分類方法で記述する。亜科単位の分類を形態や生態だけで分けるのは難しく、研究者によって諸説あったが、近年分子生物学的な手法の採用により分類が再編されつつある。
マツ亜科 Subfamily Pinoideae
[編集]以下の1属を含む単型(モノタイプ)
- マツ属 (学名Pinus)
- 枝は長枝と短枝があり、葉は短枝からのみ延びる。長枝は鱗片葉と呼ばれる特殊な葉で覆われるが、カラマツ属と違い普通の葉はつけない。葉は気孔が集まった気孔帯(英:stomatal bands)は、葉の裏面、もしくは両面にできる。球果の鱗片には突起(英:umbo)を持ち、成熟には通常で受粉後2年、ごく一部に3年かかるものもある。球果は樹上で分解せず、そのまま落下する。種子はresin vesicle(和名不明)という構造を持たない[9]。
- マツ科の中では最も広い分布範囲を持ち、繁栄しているグループで、北極圏から赤道直下までの北半球全域、旧大陸と新大陸のいずれにも合計110種程度が分布。特に新大陸のアメリカ合衆国東南部と西部山岳地帯の2地域では今も旺盛な進化が続いているとみられており種類が多い。いずれも常緑樹。高木が多いが灌木状になる高山種など各地の状況に適応した形態を見せる。
-
 マツ属の樹形の例 Pinus densiflora
マツ属の樹形の例 Pinus densiflora -
 マツ属の枝は鱗片状の葉で覆われるPinus pungens
マツ属の枝は鱗片状の葉で覆われるPinus pungens -
 マツの葉の断面の顕微鏡写真。気孔は両面にある
マツの葉の断面の顕微鏡写真。気孔は両面にある -
 マツ属の雄花
マツ属の雄花 -
 マツ属の雌花
マツ属の雌花 -
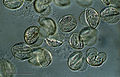 マツ属花粉の光学顕微鏡写真
マツ属花粉の光学顕微鏡写真 -
 マツ属の花粉の電子顕微鏡写真 Pinus mugo
マツ属の花粉の電子顕微鏡写真 Pinus mugo -
 二針葉松Pinus nigraの球果の鱗片の先端に発達する突起(英:umbo)
二針葉松Pinus nigraの球果の鱗片の先端に発達する突起(英:umbo) -
 五針葉松Pinus strobusの若い球果に見える突起(茶色の部分)
五針葉松Pinus strobusの若い球果に見える突起(茶色の部分) -
 マツ属の種子
マツ属の種子 -
 マツ属の発芽
マツ属の発芽


.jpg)


_Pine_tree.jpg)




_14d_1.jpg)
トウヒ亜科 Subfaimily Piceoideae
[編集]以下の1属を含む単型。
- トウヒ属 (学名:Picea)
- 球果の鱗片には突起を持たず、受粉後1年以内に成熟する。枝は長枝と短枝の区別はなく、葉は長枝につける。葉の付け根の枝に葉枕(ようちん、英:pulvinus)と呼ばれる構造が発達し、枝の凹凸が著しい。葉の気孔帯は裏面もしくは両面にできる。種子はresin vesicleを持たない。
- 長枝しか持たず常緑樹であることなどからモミ亜科に置く説もあった。
-
 トウヒ属の樹形の例 Picea abies
トウヒ属の樹形の例 Picea abies -
 トウヒ属は葉を長枝につける。
トウヒ属は葉を長枝につける。 -
 トウヒ属の枝。葉枕という構造でゴツゴツしている
トウヒ属の枝。葉枕という構造でゴツゴツしている -
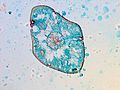 トウヒ属の葉の断面の顕微鏡写真
トウヒ属の葉の断面の顕微鏡写真 -
 トウヒ属の雄花
トウヒ属の雄花 -
 トウヒ属の雌花
トウヒ属の雌花 -
 トウヒ属の球果の鱗片には突起がない Picea abies
トウヒ属の球果の鱗片には突起がない Picea abies -
 トウヒ属の発芽
トウヒ属の発芽

_(34071357320).jpg)



.jpg)


カラマツ亜科 Subfamily Laricoideae
[編集]以下の3属を含む。球果の鱗片には突起を持たず、受粉後1年以内に成熟する。葉の気孔帯は裏面にのみでき、位置は師部よりも下側である。
- カラマツ属 (学名:Larix)
- 旧大陸、新大陸の北部や山岳地帯を中心とした冷涼な地域に10種程度が分布する。針葉樹では珍しく落葉しいずれも高木。一般には陽樹とされる。 長枝と短枝が発達する。葉は短枝の先にまとまってつくが、長枝であっても枝先端の若いものに限り葉をつける。球果は樹上で分解せず、そのまま落下する。
-
 黄葉するカラマツの仲間。この後落葉する。
黄葉するカラマツの仲間。この後落葉する。 -
 カラマツ属の枝には長枝と短枝があり、葉は原則として短枝につく
カラマツ属の枝には長枝と短枝があり、葉は原則として短枝につく -
 カラマツ属の展開した短枝の葉
カラマツ属の展開した短枝の葉 -
 カラマツ属の葉のイラスト。枝先の若い長枝に限り葉をつけており、短枝の葉と違うのがわかる。
カラマツ属の葉のイラスト。枝先の若い長枝に限り葉をつけており、短枝の葉と違うのがわかる。 -
 カラマツ属の雌花
カラマツ属の雌花 -
 カラマツ属の花粉
カラマツ属の花粉 -
 カラマツ属の球果には突起がない
カラマツ属の球果には突起がない -
 カラマツ属の発芽
カラマツ属の発芽



.jpg)
,_Corea_Heath,_Gouldsboro,_Maine.jpg)

_(19912935221).jpg)

- ギンサン属 (和名は仮称、学名Cathaya)
-
 ギンサン属の樹形の例
ギンサン属の樹形の例 -
 ギンサン属の葉
ギンサン属の葉 -
 ギンサン属の発芽
ギンサン属の発芽



- トガサワラ属(学名:Pseudotsuga)
- 学名は偽のツガという意味。ツガは日本語の栂に由来しており、後述のTsuga属の学名にもなっている。ツガ属と異なり葉の付け根に葉枕はない。枝は長枝しか持たない。また、球果から苞鱗と呼ばれる髭のようなものが発達し飛び出すのが特徴。球果は樹上で分解せず、そのまま落下する。
- 長枝しか持たず常緑樹であることなどからモミ亜科に置く説もあった。
-
 トガサワラ属の樹形の例
トガサワラ属の樹形の例 -
 トガサワラ属の枝の様子
トガサワラ属の枝の様子 -
 トガサワラ属の若い球果。苞鱗が飛び出している
トガサワラ属の若い球果。苞鱗が飛び出している -
 トガサワラ属の球果を下から見た様子 Pseudotsuga
トガサワラ属の球果を下から見た様子 Pseudotsuga -
 トガサワラ属の種子
トガサワラ属の種子 -
 トガサワラ属の発芽
トガサワラ属の発芽




_Franco.jpg)

モミ亜科 Subfamily Abietoideae
[編集]以下の6属を含む。球果の鱗片には突起を持たず、受粉後1年以内に成熟する。葉の気孔帯は裏面にのみでき、師部よりも下側である。球果には突起を持たない。種子はresin vesicleを持つ。系統学的にはほかの亜科とは早く分化したものとみられることが多い。
- モミ属 (学名:Abies)
- いずれも常緑の高木である。枝に長枝と短枝の区別はなく、葉は長枝から直接生え、雄花は枝に群生する。球果は枝から直立し樹上で分解しながら種子を散布する。
-
 モミ属の樹形の例
モミ属の樹形の例 -
 モミ属の葉の付け根部分には葉枕はない Abies alba
モミ属の葉の付け根部分には葉枕はない Abies alba -
 モミ属の葉の顕微鏡写真。気孔と師部の位置がわかる
モミ属の葉の顕微鏡写真。気孔と師部の位置がわかる -
 モミ属の雄花
モミ属の雄花 -
 モミ属の雌花
モミ属の雌花 -
 モミ属の球果は樹上に直立するAbies alba
モミ属の球果は樹上に直立するAbies alba -
 モミ属の球果は鱗片ごと分解し種を飛ばす
モミ属の球果は鱗片ごと分解し種を飛ばす -
 樹上で分解中のモミ属の球果
樹上で分解中のモミ属の球果 -
 モミ属の球果の鱗片と種子
モミ属の球果の鱗片と種子 -
 モミ属の発芽
モミ属の発芽









- ヒマラヤスギ属(学名:Cedrus)
- 常緑。枝はマツ属やカラマツ属と同じく枝は長枝と短枝を持ち、葉は短枝から生えるが若い長枝にも葉をつける。球果はモミ属と同じく枝から直立し樹上で分解しながら種子を散布する。
- 長枝と短枝を持つことからマツ亜科やカラマツ亜科に置く説もあった。いずれもユーラシア地域に分布し、地中海沿岸を中心に3種、1種だけヒマラヤ周辺に隔離分布する小グループ。
-
 ヒマラヤスギ属の樹形の例 Cedrus deodata
ヒマラヤスギ属の樹形の例 Cedrus deodata -
 ヒマラヤスギ属の枝には長枝と短枝があり、葉は短枝から出る Cedrus atlantica
ヒマラヤスギ属の枝には長枝と短枝があり、葉は短枝から出る Cedrus atlantica -
 ヒマラヤスギ属の雄花
ヒマラヤスギ属の雄花 -
 ヒマラヤスギ属の雌花
ヒマラヤスギ属の雌花 -
 ヒマラヤスギ属の球果はモミ属と同じく樹上に直立するCedrus deodata
ヒマラヤスギ属の球果はモミ属と同じく樹上に直立するCedrus deodata -
 ヒマラヤスギ属の球果の鱗片
ヒマラヤスギ属の球果の鱗片 -
 ヒマラヤスギ属の発芽
ヒマラヤスギ属の発芽





_2016-02-07_N43%C2%B031%27_E16%C2%B046%27.jpg)
_1month.jpg)
- ユサン属 (学名:Keteleeria)
- 中国からベトナムにかけて3種が分布する。球果は樹上に直立するが、樹上では分解せず、そのまま落下する。
-
 ユサン属の樹形の例
ユサン属の樹形の例 -
 ユサン属の葉
ユサン属の葉 -
 ユサン属の雄花
ユサン属の雄花 -
 ユサン属の球果
ユサン属の球果 -
 ユサン属の種子
ユサン属の種子





- ノトツガ属(和名は仮称 学名:Nothotsuga)
- 中国に一属一種 Nothotsuga longibracteata(和名未定)のみが生息する単型。枝は長枝と短枝を持つ。ツガ属の中でもっとも原始的な形質を持つものとして、Tsuga longibracteataとされることもある[11]。
-
 ノトツガ属の葉
ノトツガ属の葉 -
 ノトツガ属の球果
ノトツガ属の球果


- イヌカラマツ属(学名:Pseudolarix)
- 現存種はイヌカラマツ(Pseudolarix amabiris)のみの単型である。学名は偽のカラマツの意味。カラマツと同じく枝は長枝と短枝があり、葉は短枝に付くが、若い長枝にもつく。落葉樹である点もカラマツと共通している。球果はモミ属やヒマラヤスギ属と同じく樹上で分解しながら種子を散布する。
- 長枝と短枝の区別があり、落葉樹であることなどからカラマツ亜科に置く説もあった。
-
 イヌカラマツ属の樹形
イヌカラマツ属の樹形 -
 イヌカラマツ属の若い長枝。葉が生えている。
イヌカラマツ属の若い長枝。葉が生えている。 -
 イヌカラマツ属の短枝に生える葉
イヌカラマツ属の短枝に生える葉 -
イヌカラマツ属の雄花
-
 イヌカラマツ属の雌花
イヌカラマツ属の雌花 -
 黄葉するイヌカラマツ属
黄葉するイヌカラマツ属





- ツガ属(学名:Tsuga )
- 学名Tsugaは日本語の栂に由来する。枝は長枝のみを持つ。
-
 ツガ属の樹形の例
ツガ属の樹形の例 -
 ツガ属の枝は葉枕が発達する Tsuga sieboldii
ツガ属の枝は葉枕が発達する Tsuga sieboldii -
 ツガ属の葉は長いものと短いものが混在する。葉柄がある
ツガ属の葉は長いものと短いものが混在する。葉柄がある -
 ツガ属の球果に突起はない
ツガ属の球果に突起はない



_(31874286981).jpg)
脚注
[編集]- ^ 谷口武士 (2011) 菌根菌との相互作用が作り出す森林の種多様性(<特集>菌類・植食者との相互作用が作り出す森林の種多様性). 日本生態学会誌61(3), p311-318. doi:10.18960/seitai.61.3_311
- ^ 深澤遊・九石太樹・清和研二 (2013) 境界の地下はどうなっているのか : 菌根菌群集と実生更新との関係(<特集>森林の"境目"の生態的プロセスを探る). 日本生態学会誌63(2), p239-249. doi:10.18960/seitai.63.2_239
- ^ 岡部宏秋,(1994) 外生菌根菌の生活様式(共生土壌菌類と植物の生育). 土と微生物24, p15-24.doi:10.18946/jssm.44.0_15
- ^ 菊地淳一 (1999) 森林生態系における外生菌根の生態と応用 (<特集>生態系における菌根共生). 日本生態学会誌49(2), p133-138. doi:10.18960/seitai.49.2_133
- ^ 宝月岱造 (2010)外生菌根菌ネットワークの構造と機能(特別講演). 土と微生物64(2), p57-63. doi:10.18946/jssm.64.2_57
- ^ 東樹宏和. (2015) 土壌真菌群集と植物のネットワーク解析 : 土壌管理への展望. 土と微生物69(1), p7-9. doi:10.18946/jssm.69.1_7
- ^ a b c 清水邦義・松本清・大貫宏一朗「睡眠誘導効果をもつ(-)-酢酸ボルニルの睡眠促進効果ならびに肌質改善効果」『コスメトロジー研究報告』第24巻、コスメトロジー研究振興財団、2016年、181-189頁、NAID 40020955101。
- ^ 藤崎洋子・島瀬初美・五十嵐隆雄・山田康子・佐藤尚. 1976. 花粉症の研究 Ⅳマツ属花粉症. アレルギー25(9) pp668-677. doi:10.15036/arerugi.25.668
- ^ David Kolotelo. 2005. Cone and Seed Imprpvement BCMoF tree centre -Resin Vesicles in Conifer Seeds -
- ^ Ran, Jin-Hua; Shen, Ting-Ting; Wu, Hui; Gong, Xun; Wang, Xiao-Quan 2018. Phylogeny and evolutionary history of Pinaceae updated by transcriptomic analysis. Molecular Phylogenetics and Evolution. 129: 106–116.
- ^ 清水建美. 1990, 針葉樹の分類・地理、特に2、3の亜高山帯の属について その1. 植生史研究(6) 25-30.
外部リンク
[編集]- Arboretum de Villardebelle フランスの樹木園
- Gymnosperm Database – マツ科 Pinaceae
- Pinus in USDA Plants Database アメリカ合衆国農務省の植物データベース
この項目は、植物に関連した書きかけの項目です。この項目を加筆・訂正などしてくださる協力者を求めています(プロジェクト:植物/Portal:植物)。 |